Мітоз
Міто́з (рідше каріокінез або непрямий поділ) (від дав.-гр. μίτος — нитка) — найпоширеніший спосіб поділу ядер клітин. Під час мітозу два ідентичні набори хромосом, утворені внаслідок реплікації ДНК, розподіляються між двома новими ядрами. Мітозом діляться ядра всіх клітин багатоклітинних організмів, крім гамет[1].


Зазвичай після ядра ділиться й цитоплазма (процес цитокінезу), і в результаті материнська клітина повністю ділиться на дві дочірні. Проте в низці випадків цитокінез не відбувається, і утворені ядра лишаються в одній клітині. Прикладами можуть бути деякі клітини ембріона плодової мухи, гепатоцити та кардіоміоцити ссавців. Якщо ж цитокінез відбувається, він починається в анафазі й закінчується під час або одразу після телофази[1].
Мітоз і цитокінез складають M-фазу клітинного циклу, завдяки якій зберігається сталий каріотип багатоклітинного організму і виду в цілому та відбувається збільшення кількості клітин.
У різних груп еукаріот мітоз відбувається дещо по-різному. Прокаріотичні клітини діляться бінарним поділом[en], що відмінний від мітозу.
Мітоз — лише одна з частин клітинного циклу, але під час мітозу відбуваються значні клітинні перебудови, тому його ділять на п'ять фаз: профаза, прометафаза, метафаза, анафаза і телофаза. Іноді прометафазу не виокремлюють.
- У профазі відбувається конденсація хромосом, клітинний центр ділиться і продукти його поділу розходяться до полюсів ядра, руйнується ядерна оболонка, утворюється веретено поділу.
- У прометафазі каріоплазма і цитоплазма змішуються, хромосоми ще більше ущільнюються, на центромерах утворюються кінетохори, до них приєднуються мікротрубочки веретена поділу.
- У метафазі хромосоми розташовуються на екваторі (на рівній відстані від полюсів ядра) в одній площині, утворюючи так звану метафазну пластинку.
- У анафазі з'єднання подвоєних ідентичних хромосом у районі центромери руйнується, хроматиди розділяються і розходяться до різних полюсів клітини.
- У телофазі руйнується веретено поділу, утворюється ядерна оболонка навколо дочірніх ядер.
Подвоєння хромосом відбувається ще в інтерфазі, що не є частиною M-фази. В результаті цього в мітоз хромосоми вступають вже подвоєними, такими, що нагадують букву «X» (ідентичні копії материнської хромосоми, сполучені одна з одною в області центромери) (див. п. Мітотична хромосома)
Фази мітозу
ред.
Тривалість мітозу залежить від виду клітин і в середньому становить 1-2 години. Процес залежить і від умов зовнішнього середовища (температури, світлового режиму й інших показників).
Профаза
ред.
Профаза є першою стадією мітозу. Під час профази конденсуються хромосоми та формується веретено поділу. У деяких організмів, зокрема тварин та рослин, руйнується ядерна оболонка (відкритий мітоз)[2]. Якщо вона не руйнується (закритий мітоз, як у більшості грибів), веретено поділу формується всередині неї[3][4].
- Руйнування ядерної оболонки
Ядерна мембрана двошарова. Внутрішній шар прилягає до хроматину, а зовнішній переходить в ендоплазматичний ретикулум. Ядерну оболонку пронизують ядерні пори та підтримує спеціалізована структура — ламіна, що складається з білків ламінів[en][3].
Руйнування ядерної оболонки починається в профазі. Фосфорилювання білків внутрішньої мембрани, ламінів та деяких інших білків циклін-залежною протеїнкіназою 1 (CDK1)[5] та іншими протеїнкіназами, зокрема Aurora B, порушує зв'язок хроматину та ядерної оболонки. Інвагінації та утворення дірок (fenestration) в ядерній оболонці тісно пов'язані з активністю динеїнових молекулярних моторів, що приєднані до ядерних пор. Динеїни здатні до активного переміщення вздовж мікротрубочок і рухаються по астральним мікротрубочкам веретена поділу. Таким чином одночасно відбувається розміщення веретена поділу по полюсам клітини і формування дірок в ядерній оболонці.[3]
Прометафаза
ред.
Під час прометафази закінчується конденсація хромосом та руйнування ядерної оболонки. Веретено поділу розміщується на полюсах клітини. Від центросом полімеризуються та ростуть мікротрубочки всіх типів (див. п. Веретено поділу). Кінетохор деяких хромосом взаємодіє з кінетохорними мікротрубочками веретена поділу за допомогою білків динеїну та кінезину CENP-E[6], що допомагають латерально приєднатися до мікротрубочки, та білку NDC80[7], що формує стабільний зв'язок з плюс-кінцем мікротрубочки.[8]
Після приєднання до кінетохору однієї мікротрубочки до нього приєднуються інші мікротрубочки, причому їхня полімеризація може не залежати від центросоми. Так, до існуючої кінетохорної мікротрубочки може приєднатися комплекс білків Augmin разом з γTuRC. Від цього комплексу може сформуватися нова мікротрубочка, яка буде йти майже паралельно до існуючої і в тому ж просторовому напрямку, тобто плюс-кінцем до кінетохору. Таким чином може швидко сформуватися к-фібрила (кінетохорна фібрила): пучок мікротрубочок, який необхідний для руху хромосом під час анафази.[3]
Важливим моментом під час прометафази є початок активації контрольної точки веретена поділу[en], яка не дозволить клітині вхід в анафазу за відсутності приєднання всіх кінетохорів до мікротрубочок (див. Контроль мітозу).[8]
Метафаза
ред.
Під час метафази хромосоми розміщуються вздовж екватора клітини. Таке розміщення хромосом викликано з'єднанням обох кінетохорів однієї хромосоми до мікротрубочок протилежних центросом. Клітина перебуває у метафазі довгий час, допоки кожна хромосома не буде з'єднана з обома центросомами з обох полюсів.

Правильний процес під'єднання кінетохорів до мікротрубочок від різних полюсів забезпечується методом спроб і помилок, таке правильне з'єднання має велике тягове зусилля: хромосому тягнуть кінезинові білки до різних полюсів клітини і лише з'єднання в області центромери не дає сестринським хроматидам розійтися. Таке з'єднання є стабільним. При неправильному контакті, наприклад контакті обох кінетохорів з мікротрубочками одного полюсу, цього тягового зусилля не виникає, таке з'єднання є нестабільним і швидко руйнується, тоді кінетохор знову з'єднується з мікротрубочками і такий процес триває, доки не утвориться стабільне з'єднання. Причина стабільності з'єднання в основному лежить у кіназній активності білку Aurora B, який фосфорилює комплекс приєднання мікротрубочок до веретена поділу, що призводить до розриву контакту. Одна з моделей, яка пояснює цей механізм, називається «собача прив'язь» і полягає в тому, що Aurora B приєднана до внутрішнього кінетохору за допомогою білку INCENP[9] і може фосфорилювати лише ті білки, що знаходяться в радіусі рухливості INCENP, на кшталт прив'язі собаки[10]. Таким чином, при неправильному контакті напруги між кінетохором і мікротрубочками немає і плюс-кінці та білкові комплекси зовнішнього кінетохору, що контактують з мікротрубочок, лежать в зоні досягнення Aurora B, що їх фосфорилює та руйнує контакт. При контакті хромосоми з мікротрубочками обох полюсів виникає тягова сила, відстань між внутрішнім і зовнішнім кінетохором збільшується, Aurora B вже не досягає до комплексу з'єднання з мікротрубочками і нездатний його фосфорилювати. Тому такий контакт залишається[10].
Таким чином, кожна хромосома перетягується в протилежних напрямках до обох полюсів клітини і мігрує в центр (див. п. Рух хромосом), оскільки пряма лінія є геометрично найкоротшою відстанню між двома точками. Хромосоми розміщуються вздовж екватора, формуючи метафазну пластинку.
Анафаза
ред.
Під час анафази контакт між сестринськими хроматидами розривається, і вони рухаються до протилежних полюсів клітини. Перехід від метафази до профази можливий лише при проходженні контрольної точки завдяки діяльності комплексу APC (див. п. Контроль мітозу).
Анафаза ділиться на дві частини:[1]
- A-анафаза, перша частина анафази. Виникає при різкому розриванні контактів між сестринськими хроматидами. До цього моменту мікротрубочки тягнули кожен з кінетохорів у бік полюсу, але когезиновий контакт запобігав розходженню хроматид. При руйнуванні контакту дочірні хромосоми різко починають рухатися у бік полюсу. У цій стадії хроматин набуває найбільшої щільності під час мітозу[11]
- B-анафаза, пізня анафаза. Відбувається коли дочірні хромосоми вже трохи відійшли до полюсів. Тоді починають рухатися у протилежні боки полюси веретена поділу.
Рух хромосом
ред.Хромосоми здійснюють значні переміщення в клітині під час мітозу навіть до стадії анафази: під час метафази вони вибудовуються в площині екватора. Рух хромосом та витягнення веретена поділу здійснюються завдяки трьом механізмам.[1]
- Перший призводить до руху дочірньої хромосоми до полюса веретена поділу. Цей механізм не до кінця з'ясований, але він не залежить від моторних білків та не потребує гідролізу АТФ. Він досягається за допомогою деполімеризації плюс-кінця мікротрубочки.
- Другий механізм полягає в русі мікротрубочок та з'єднаних з ними хромосом до полюса клітини, імовірно через деполімеризацію мінус-кінця мікротрубочок. При цьому плюс-кінці мікротрубочок, навпаки, полімеризуються. Цей процес називається «потік мікротрубочок у напрямку до полюса» (англ. poleward microtubule flux)[12].
- І третій механізм, який не є активним в анафазі, проте дуже потрібний в прометафазі та метафазі, полягає в русі хромосом від полюса клітини. Це забезпечується завдяки активності моторних білків: кінезину 4 та кінезину 10, які здатні рухатися по мікротрубочці у напрямку від мінус- до плюс-кінця.[1]
Телофаза
ред.
Телофаза є завершальною стадією мітозу, за нею слідує цитокінез. Під час телофази навколо дочірніх хромосом, які розміщені на протилежних полюсах клітини, формується ядро, хромосоми займають свої інтерфазні позиції (хромосомні території), еухроматин декомпактизується, що дозволяє зчитувати гени. Активною є РНК-полімераза I[en], що транскрибує рРНК і формуються ядерця навколо ядерцевих організаторів.[13] Також під час телофази руйнується веретено поділу.[1]
- Формування ядерної оболонки
Інактивація комплексу ЦЗК1–циклін B ініціює формування ядерної мембрани. Частини внутрішньої ядерної мембрани повертаються від ендоплазматичного ретикулуму і з'єднуються з хроматином. При цьому хроматин також зазнає певних змін; так, від нього від'єднується кіназа Aurora B (див. п. мітотична хромосома). Злиття частин ядерної мембрани відбувається одночасно з утворенням ядерних пор, таким чином уникається формування закритого ядра без пор.[3]
Формування ядерної мембрани повинно відбутися таким чином, що всі хромосоми опиняться всередині ядра, а органели, навпаки, туди не потраплять. Цьому процесу допомагає щільна (щільніша, ніж у метафазі) упаковка хромосом та присутність веретена поділу на перших стадіях телофази: хромосоми розміщені щільною купкою, а навколо них формується ядерна оболонка. Тим не менш у цьому процесі можливі помилки, в результаті яких губиться хромосома чи формуються мікроядра.[3]
Клітинний цикл, що межує з мітозом
ред.Мітоз займає лише частину клітинного циклу. Проте для проходження мітозу стадії до нього також повинні пройти успішно. Так, передує мітозу G2-стадія інтерфази, а закінчується мітоз цитокінезом, і переходить в G1-фазу інтерфази.
Клітинна анатомія мітозу
ред.
Веретено поділу
ред.Під час мітозу вибудувані під час метафази в екваторіальній площині хромосоми повинні розійтися на два полюси клітини, причому розділитися правильно на дві частини. Це надзвичайно складний процес, в якому беруть участь багато білків, але основною структурою, яка його забезпечує, є веретено поділу.

Веретено поділу складається з декількох типів мікротрубочок та центросом.
- Мікротрубочки
Це довгі фібрили білкового комплексу, який складається з димерів α- та β-тубуліну, які, спірально накручуючись, складають структуру, схожу на трубу, окружністю 13 αβ-тубулінових димерів. Така структура несиметрична та має плюс- та мінус-кінці: перший містить β-субодиницю, останній α-субодиницю[14]. Також на мінус-кінці мікротрубочки веретена поділу є γ-тубулін. Мікротрубочки — нестабільні структури і постійно руйнуються та синтезуються з плюс-кінця (мінус-кінець закріплений γ-тубуліном). Мікротрубочки веретена поділу формуються з центра організації мікротрубочок (ЦОМТ), в основі якого лежить γ-тубулін γTuRCs (англ. γ-tubulin ring complex).
До складу веретена поділу входять:[15]
- кінітохорні мікротрубочки (англ. kinetochore microtubule, K-MT), плюс-кінці яких йдуть до кінетохору;
- астральні мікротрубочки (англ. astral microtubule, A-MT), плюс-кінці яких йдуть до цитоскелета, радіально від полюса, допомагаючи закріпитися веретену поділу у клітині та виконуючи роль під час цитокінезу;
- не-кінетохорні мікротрубочки (англ. non-kinetochore microtubule, nK-MTs) чи міжполюсні мікротрубочки. Доходять приблизно до центру клітини, перетинаючись з не-кінетохорними мікротрубочками протилежного полюсу.
Мінус-кінці всіх цих мікротрубочок розташовані на полюсі.[15]
До одного кінетохору зазвичай приєднується не одна, а 20-40 мікротрубочок, формуючи кінетохорну фібрилу (к-фібрилу)[16].
- Центральна лінія веретена поділу

Починаючи з пізньої анафази і до цитокінезу не-кінетохорні мікротрубочки протилежних полюсів перетинаються і формують у центрі клітини спеціальну структуру — центральну лінію веретена поділу (англ. spindle midzone).[17] Ця структура потрібна для розходження полюсів веретена поділу під час пізньої анафази і телофази, і вона формує основу для актинової перетяжки, що ділить цитоплазму клітини надвоє під час цитокінезу.
У дріжджів основою центральної лінії веретена поділу є білок Ase1[18][19].
- Центросома

У тваринних клітинах центросоми є центрами на полюсах, де збираються мікротрубочки веретена поділу. Центросоми складаються з центріолей. Навколо центріолей розміщена перицентріолярна речовина (англ. pericentriolar material, PCM)[20].
У ранній інтерфазі в клітині одна центросома — та, що дісталася з половини веретена попереднього поділу. Проте перед наступним поділом клітини центросома також подвоюється. Цей процес починається в S-фазі.[20][21]
Центріоль, яка лежить в центрі центросоми, утворює дочірню центріоль таким чином, що вони лежать під кутом 90°. Центріолі хребетних тварин містять дев'ять триплетів мікротрубочок, що радіально-симетрично розташовані навколо колоподібної структури (англ. cartwheel) та оточені додатковими білками. Довжина центріолі хребетних ~300-500 нм. Центріолі D. melanogaster простіші, мають менше білків, замість триплетів мікротрубочок мають диплети, також вони коротші, довжиною ~150 нм.[20] При подвоєнні центріолі в S-фазі центральна колоподібна структура руйнується, залишаючи приєднані один з одним дві центріолі[21].
Центросоми не є необхідними для поділу клітини і правильної організації мікротрубочок, але їхня відсутність запускає p53-залежну заборону клітинної проліферації в людини та миші, але не у плодових мух. Наприклад, у лінії мух, мутантної за геном Sas4 (англ. Spindle assembly abnormal 4 orthologue)[22], центросоми не формуються. Ранній ембріогенез у таких організмів проходить погано, проте якщо їм експериментально надати невелику кількість білку Sas4, ембріогенез проходить нормально, центріолі швидко губляться, мухи народжуються фенотипово нормальні. Проте за відсутності центріолей клітини не можуть формувати джгутики, для яких вони необхідні.[20]
Мітотична хромосома
ред.Під час мітозу хромосоми повинні знаходитися в конденсованому стані — інтерфазний («рихлий») хроматин, який дозволяє зчитування генів з ДНК, фізично не дозволяє значні рухи хромосоми в клітині, адже кожна молекула ДНК дуже довга. Під час профази хромосоми конденсуються повністю. Під час ранньої анафази хроматин набуває найбільшого ущільнення впродовж мітозу, тоді як під час пізньої анафази та впродовж телофази хромосоми починають зменшувати свою щільність упаковування. [11] Проте хроматин не упакований настільки щільно, що він недоступний до певних факторів транскрипції, нуклеаз чи вставок транспозонів.[23] Так, в експериментах на сайти чутливості до ДНКази I[en] та ATAC-секвенування[en] встановлено, що доступність мітотичного хроматину не сильно відрізняється від доступності хроматину в інших фазах клітинного циклу для транспозонів чи до розрізання DNase1-нуклеазою.[23] Через це теоретична ідея, що мітотична ДНК занадто щільно спакована для того, щоб до неї дістався транскрипційний апарат, не вірна.[23]
Мітотичні хромосоми мають характерну X-подібну форму. Така форма притаманна лише мітотичній хромосомі, причому до стадії анафази, коли з'єднання в області центромери руйнується і сестринські хроматиди розходяться до полюсів. Таку форму забезпечують конденсованість хроматину та з'єднання сестринських хроматид в області центромери. Під час подвоєння ДНК в період S-фази дочірні копії ДНК з'єднуються за допомогою білку когезину[24] вздовж всієї довжини ДНК в період фази G2, до початку мітозу[25]. Проте при вступі в мітоз когезин фосфорилюється протеїнкіназами CDK1, PLK1[26] та Aurora B. Білок Wapl (англ. Wings apart-like protein homolog)[27] від'єднує когезин від хромосом, причому лише в області плечей хромосоми, а не від центромери, де когезин знаходиться в дефосфорильованому стані завдяки активності білка Sgo1[28]. Таким чином когезин при вступі в мітоз залишається лише в області центромери, і хромосоми мають X-подібний вигляд (чи V-подібний, якщо це акроцентричні хромосоми, у яких центромера знаходиться з краю, наприклад Y-хромосома)[25].
- Центромера
Центромери — це специфічні ділянки хромосом, де формується кінетохор під час мітозу і куди приєднуються мікротрубочки веретена поділу. Типова хромосома має лише одну центромеру. Проте формування та встановлення центромери не прямо залежить від послідовності ДНК. У багатьох організмів ДНК що формують центромери містять повторювані ділянки[en], такі як α-сателіти, проте позиціонування та формування центромери залежить від епігенетичних механізмів. Так, гістон H3 замінюється в ділянці центромери на специфічний гістоновий варіант CENPA. Також існують дані про незначний дрейф локації центромери після багатьох поділів клітини, особливо в тих випадках коли в центромери немає білків CENPU та CENPS[29]
Конденсація та деконденсація хромосом
ред.Конденсація хроматину для проходження мітозу проходить під впливом багатьох білкових комплексів та факторів, модифікацій гістонових хвостів тощо.[11] При цьому сама по собі довжина молекули ДНК в хромосомах викликає питання, як хромосоми можуть перейти від рихлого до конденсованого стану окремих молекул без того, аби заплутатися між собою.[30] Для пояснення цієї топологічної проблеми часто використовують порівняння локшини чи навушників: взяти виделкою окремо довгу локшину з тарілки спагеті важко.[31][32][33]
Кінетохор
ред.Кінетохор — ділянка хромосоми в області центромери. До кінетохору кріпляться мікротрубочки веретена поділу, що рухають хромосоми під час анафази[34].
Кінетохор поділяється на[14]
- внутрішній кінетохор: частина кінетохору, що розміщена біля хроматину;
- проміжний кінетохор;
- зовнішній кінетохор: взаємодіє з мікротрубочками. Має товщину 50-60 нм.
Треба розуміти, що таке розділення не є дуже чітким, однак дані шари формуються різними групами білків. Також кінетохору притаманна тимчасова структура — фіброзна зона — при відсутності контакту з мікротрубочками.
Внутрішній кінетохор формується білками CENP-A, що є варіантами гістону H3[en]. Ці білки формують конститутивну з'єднану з центромерою мережею (англ. constitutive centromere-associated network, CCAN), що названа так тому що ці білки взаємодіють з центромерою впродовж всього клітинного циклу, а не лише у певних фазах підготовки до мітозу.[35]
Починаючи з фази G2 клітинного циклу на внутрішньому кінетохорі будується комплекс зовнішнього кінетохору, який в основному складається з білків KMN, що формується білковими комплексами KNL1, MIS12 та NDC80.[35]
Загальні зміни в клітині
ред.Крім наявності таких специфічних структур, як веретено поділу, клітина зазнає значних загальних змін при вході в мітоз з G2-фази. Це включає округлення загальної форми клітини, зміни цитоскелету та органел, зникнення ядерної оболонки.[2]
- Форма клітини

В інтерфазі клітини мають різну форму залежно від своїх функцій, проте при вступі в мітоз вони стають округлими. Це спостерігається й на клітинах у культурі in vitro, які зазвичай плоскі з випуклістю в центрі. Під мембраною клітини формується щільний шар цитоскелету — клітинний кортекс[en] — який складається з актинових філаментів, білків, що з ними з'єднані, та міозинових моторних білків[2].
Контроль мітозу
ред.Під час мітозу відбувається серія клітинних перебудов та рухів різних органел, а в результаті мітозу повинні утворитися дві максимально однакові дочірні клітини, генетичний матеріал яких поділений навпіл. Це дуже складний процес і його контроль відбувається на декількох рівнях і в декілька етапів.
Для правильного проходження мітозу повинні бути пройдені контрольні точки клітинного циклу[en] — певні моменти в стадіях мітозу, при яких повинні бути виконані серії вимог і лише в такому разі настає подальша стадія. У мітозу контрольних точок є дві:[1]
- вхід в мітоз зі стадії G2; при проходженні цієї точки відбуваються перші стадії: профаза, прометафаза і метафаза;
- система контролю збірки веретена поділу (англ. spindle assembly checkpoint, SAC) яка перевіряє приєднання кінетохорів кожної сестринської хромосоми до мікротрубочок веретена поділу, не дозволяючи перехід в анафазу до того, як всі хромосоми не будуть приєднанні до веретена поділу належним чином.
Система контролю збірки веретена поділу
ред..svg)
Система контролю збірки веретена поділу, яку іноді називають мітотичною системою контролю, — це запобіжний механізм, який протидіє некоректному розподіленню та втраті хромосом під час мітозу. Ця система затримує клітину в метафазі доки всі хромосоми не буде правильно приєднано до веретена поділу. Час затримки може сягати до 20 годин у людини та декількох годин у дріжджів[36]
В системі контролю збірки веретена поділу в основі реагує на стабільність приєднання кінетохорів до мікротрубочок. Це забезпечується низкою білків та білкових комплексів:[36]
- Комплекс мітотичного контролю (англ. mitotic checkpoint complex, MCC), що інгібує дію комплексу APC/C. До нього входять білки Mad2, BubR1, Bub3 та Cdc20, причому для активності Mad2 необхідне з'єднання з Cdc20.[37]
- комплекс APC/C (англ. anaphase-promoting complex чи cyclosome) — убіквітин-лігаза, що може здійснювати убіквітинування секьюрину
- секьюрин — білок, який не дає діяти сепаразі.
Відповідно, при вимиканні MCC комплекс APC/C убіквітинує секьюрин. Секьюрин, з'єднаний з убіквітиновими залишками, стає мішенню руйнування в протеасомі. Таким чином вивільнюється сепараза, яка розщеплює білок Scc1, що призводить до руйнування когезинів в центромерній ділянці і розходження сестринських хроматид.
Варіанти мітозу
ред.
Слід зазначити, що в різних групах живих організмів мітоз протікає дещо по-різному. Описаний вище варіант мітозу називається відкритим ортомітозом (ядерна оболонка руйнується, веретено поділу пряме, оскільки продукти поділу клітинного центру розміщені на протилежних полюсах ядра). Характерний для багатоклітинних тварин, багатоклітинних рослин і ряду найпростіших[4].
- Плевромітоз
У деяких групах найпростіших продукти поділу клітинного центру в анафазі не досягають протилежних сторін ядра, внаслідок чого мікротрубочки веретена поділу розташовуються під кутом, нагадуючи букву V.
- Закритий та напівзакритий мітоз
У деяких організмів, таких як пивні дріжджі, мітоз відбувається без руйнування ядерної оболонки. В такому випадку формуються веретеноподібні полюсні тіла (англ. spindle pole bodies) — схожі на центросоми структури, що є центрами організації мікротрубочок, але формуються всередині ядра та приєднуються зсередини до його мембрани (постійно або тимчасово). При анафазі ядерна мембрана закритого мітозу змінює форму та збільшує площу, разом з сіткою ендоплазматичного ретикулюму, що оточує ядро. Це досягається завдяки активності ферменту lipin, фосфатази, що збільшує кількість фосфоліпідів, які здатні вбудуватися в мембрану ядра, збільшивши її. У деяких дріжджів, таких як Schizosaccharomyces japonicus білок lipin неактивний, мембрана ядра не збільшується, тому під час анафази при розходженні хромосом ядерна оболонка руйнується. Такий тип мітозу називається напівзакритим[3].
- Центріолярний і ацентріолярний
Клітинний центр може містити центріолі (наприклад, у тварин) або не містити їх (наприклад, у квіткових рослин).
Історія відкриття
ред.У 1874 І. Д. Чистяков описав ряд стадій (фаз) мітозу у спорах плаунів, ще не ясно представляючи собі їхню послідовність. Детальні дослідження з морфології мітозу уперше були виконані Е. Страсбургером на рослинах (1876–1879).
Мітоз у тварин вперше незалежно описали Вальтер Флеммінг, В. Шляйхер[38] і Петро Перемежко[39]. Засновник кафедри гістології, ембріології і порівняльної анатомії в Університеті Св. Володимира Петро Перемежко (1833–1893), на результатах власних досліджень, проведених у Анатомічному театрі в Києві, у 1878 році відкрив непрямий поділ тваринних клітин.[40]
Мікроскопія мітозу
ред.- Флуоресцентна мікроскопія мітозу
-
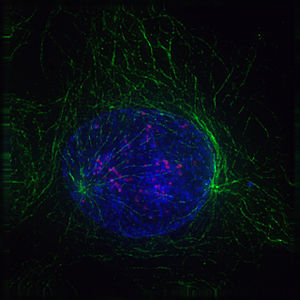 Профаза
Профаза -
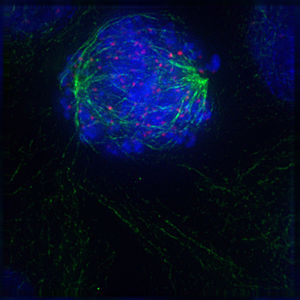 Прометафаза
Прометафаза -
 Метафаза
Метафаза -
 Анафаза
Анафаза -
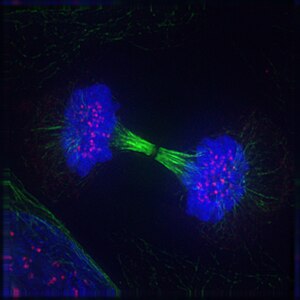 Телофаза
Телофаза






-
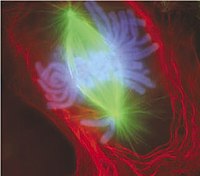 Рання анафаза мітозу в клітині нирки тритона (оптична мікрофотографія, імунофлюоресценція). Мікротрубочки зелені, хромосоми блакитні.
Рання анафаза мітозу в клітині нирки тритона (оптична мікрофотографія, імунофлюоресценція). Мікротрубочки зелені, хромосоми блакитні. -
 Конфокальна мікрофотографія мітотичних мікротрубочок (забарвлені зеленим за допомогою анти-тубуліну, кон'югованого з барвником) та ядер (блакитні). Клітини меристеми головних коренів чотириденних проростків Arabidopsis thaliana (Col-0). Довжина масштабної лінійки — 5 мкм.
Конфокальна мікрофотографія мітотичних мікротрубочок (забарвлені зеленим за допомогою анти-тубуліну, кон'югованого з барвником) та ядер (блакитні). Клітини меристеми головних коренів чотириденних проростків Arabidopsis thaliana (Col-0). Довжина масштабної лінійки — 5 мкм.


Див. також
ред.Примітки
ред.- ↑ а б в г д е ж Bruce Alberts, Alexander Johnson, Julian Lewis, David Morgan, Martin Raff Keith Roberts, and Peter Walter (2014). Molecular biology of the cell (англ.) (вид. 6). Garland Science. ISBN 9780815344322. OCLC 902708011.
- ↑ а б в Champion, Lysie; Linder, Monika I.; Kutay, Ulrike (2016). Cellular Reorganization during Mitotic Entry. Trends in Cell Biology. Т. 27, № 1. с. 26—41. doi:10.1016/j.tcb.2016.07.004. Архів оригіналу за 25 червня 2018. Процитовано 25 лютого 2017.
- ↑ а б в г д е ж Ungricht, Rosemarie; Kutay, Ulrike (2017). Mechanisms and functions of nuclear envelope remodelling. Nature Reviews Molecular Cell Biology. doi:10.1038/nrm.2016.153.
- ↑ а б Boettcher B., Barral Y. (2013). The cell biology of open and closed mitosis (PDF). Nucleus. doi:10.4161/nucl.24676.
- ↑ Циклін-залежна протеїнкіназа 1 людини в базі даних UniProt P06493
- ↑ Центромерний білок E людини в базі даних UniProt Q02224
- ↑ Білок кінетохору Ndc80 людини в базі даних UniProt O14777
- ↑ а б Trivedi, Prasad; Stukenberg, P. Todd (1 лютого 2016). A Centromere-Signaling Network Underlies the Coordination among Mitotic Events. Trends in Biochemical Sciences. Т. 41, № 2. с. 160—174. doi:10.1016/j.tibs.2015.11.002. ISSN 0968-0004. PMC 4733523. PMID 26705896. Архів оригіналу за 24 березня 2017. Процитовано 27 лютого 2017.
{{cite news}}: Обслуговування CS1: Сторінки з PMC з іншим форматом (посилання) - ↑ Білок Inner centromere protein людини в базі даних UniProt Q9NQS7
- ↑ а б Krenn, Veronica; Musacchio, Andrea (1 січня 2015). The Aurora B kinase in chromosome bi-orientation and spindle checkpoint signaling. Molecular and Cellular Oncology. с. 225. doi:10.3389/fonc.2015.00225. PMC 4607871. PMID 26528436.
{{cite news}}: Обслуговування CS1: Сторінки з PMC з іншим форматом (посилання) Обслуговування CS1: Сторінки із непозначеним DOI з безкоштовним доступом (посилання) - ↑ а б в Antonin, Wolfram; Neumann, Heinz (June 2016). Chromosome condensation and decondensation during mitosis. Current Opinion in Cell Biology. Т. 40. с. 15—22. doi:10.1016/j.ceb.2016.01.013. ISSN 1879-0410. PMID 26895139. Архів оригіналу за 23 липня 2018. Процитовано 23 березня 2018.
{{cite news}}: Cite має пустий невідомий параметр:|14=(довідка) - ↑ Akhmanova, Anna; Hoogenraad, Casper C. (16 лютого 2015). Microtubule minus-end-targeting proteins. Current biology: CB. Т. 25, № 4. с. R162—171. doi:10.1016/j.cub.2014.12.027. ISSN 1879-0445. PMID 25689915. Архів оригіналу за 26 березня 2017. Процитовано 13 березня 2017.
- ↑ McStay, Brian (15 липня 2016). Nucleolar organizer regions: genomic 'dark matter' requiring illumination. Genes & Development. 30 (14): 1598—1610. doi:10.1101/gad.283838.116. ISSN 1549-5477. PMC 4973289. PMID 27474438. Архів оригіналу за 26 березня 2017. Процитовано 13 березня 2017.
{{cite journal}}: Обслуговування CS1: Сторінки з PMC з іншим форматом (посилання) - ↑ а б А. В. Сиволоб, К. С. Афанасьєва (2012). Молекулярна організація хромосом (PDF) (укр.). К: Видавничо-поліграфічний центр «Київський університет». с. 249—293. Архів оригіналу (PDF) за 23 вересня 2015. Процитовано 25 лютого 2017.
- ↑ а б Prosser, Suzanna L.; Pelletier, Laurence (2017). Mitotic spindle assembly in animal cells: a fine balancing act. Nature Reviews Molecular Cell Biology. 18 (3): 187—201. doi:10.1038/nrm.2016.162.
- ↑ eLife digest. eLife (англ.). 2015. doi:10.7554/eLife.07635.002. ISSN 2050-084X. Архів оригіналу за 28 лютого 2017. Процитовано 27 лютого 2017.
{{cite journal}}: Обслуговування CS1: Сторінки із непозначеним DOI з безкоштовним доступом (посилання) - ↑ Mierzwa, Beata; Gerlich, Daniel W. (2014). Cytokinetic Abscission: Molecular Mechanisms and Temporal Control. Developmental Cell. 31 (5): 525—538. doi:10.1016/j.devcel.2014.11.006. Архів оригіналу за 30 червня 2018. Процитовано 6 березня 2017.
- ↑ Білок Anaphase spindle elongation protein S. cerevisiae в базі даних UniProt P50275
- ↑ Khmelinskii, Anton; Schiebel, Elmar (1 лютого 2008). Assembling the spindle midzone in the right place at the right time. Cell Cycle (Georgetown, Tex.). 7 (3): 283—286. doi:10.4161/cc.7.3.5349. ISSN 1551-4005. PMID 18235228. Архів оригіналу за 24 березня 2017. Процитовано 6 березня 2017.
- ↑ а б в г Conduit, Paul T.; Wainman, Alan; Raff, Jordan W. (2015). Centrosome function and assembly in animal cells. Nature Reviews Molecular Cell Biology. 16 (10): 611—624. doi:10.1038/nrm4062.
- ↑ а б Loncarek, Jadranka; Bettencourt-Dias, Mónica (28 грудня 2017). Building the right centriole for each cell type. The Journal of Cell Biology. doi:10.1083/jcb.201704093. ISSN 1540-8140. PMID 29284667. Архів оригіналу за 5 червня 2018. Процитовано 25 січня 2018.
- ↑ Білок Sas4 D. melanogaster в базі даних UniProt Q9VI72
- ↑ а б в Palozola, Katherine C.; Lerner, Jonathan; Zaret, Kenneth S. (2019-01). A changing paradigm of transcriptional memory propagation through mitosis. Nature Reviews. Molecular Cell Biology. Т. 20, № 1. с. 55—64. doi:10.1038/s41580-018-0077-z. ISSN 1471-0080. PMC 6557398. PMID 30420736.
- ↑ Субодиниця когезину SA-1 людини в базі даних UniProt Q8WVM7
- ↑ а б Haarhuis, Judith H. I.; Elbatsh, Ahmed M. O.; Rowland, Benjamin D. (2014). Cohesin and its regulation: on the logic of X-shaped chromosomes. Developmental Cell. Т. 31, № 1. с. 7—18. doi:10.1016/j.devcel.2014.09.010. ISSN 1878-1551. PMID 25313959. Архів оригіналу за 24 березня 2017. Процитовано 28 лютого 2017.
- ↑ Протеїнкіназа PLK1 людини в базі даних UniProt P53350
- ↑ Білок WAPL людини в базі даних UniProt Q7Z5K2
- ↑ Білок SGO1 (Shugoshin 1) людини в базі даних UniProt Q5FBB7
- ↑ Hori, Tetsuya; Kagawa, Naoko; Toyoda, Atsushi; Fujiyama, Asao; Misu, Sadahiko; Monma, Norikazu; Makino, Fumiaki; Ikeo, Kazuho; Fukagawa, Tatsuo (2 січня 2017). Constitutive centromere-associated network controls centromere drift in vertebrate cells. J Cell Biol (англ.). Т. 216, № 1. с. 101—113. doi:10.1083/jcb.201605001. ISSN 0021-9525. PMID 27940888. Архів оригіналу за 20 березня 2018. Процитовано 19 березня 2018.
{{cite news}}: Cite має пустий невідомий параметр:|30=(довідка) - ↑ Brangwynne, Clifford P.; Marko, John F. (2016/07). A sticky problem for chromosomes. Nature (англ.). Т. 535, № 7611. с. 234—235. doi:10.1038/nature18904. ISSN 1476-4687. Архів оригіналу за 19 січня 2022. Процитовано 23 березня 2018.
- ↑ L., Hartl, Daniel (2014). Essential genetics : a genomics perspective (вид. 6th ed). Burlington, MA: Jones & Bartlett Learning. с. 94. ISBN 9781449686888. OCLC 786161787.
- ↑ Mystery of the instant noodle chromosomes: Researchers evaluated benefits of placing the DNA on principle of spaghetti. ScienceDaily (англ.). Архів оригіналу за 24 березня 2018. Процитовано 23 березня 2018.
{{cite news}}: Cite має пустий невідомий параметр:|5=(довідка) - ↑ The human has 46 double chromosomes or simple chromosomes?. biology.stackexchange.com. Архів оригіналу за 24 березня 2018. Процитовано 23 березня 2018.
{{cite web}}: Cite має пустий невідомий параметр:|4=(довідка) - ↑ Musacchio, Andrea (2016). The Molecular Biology of Spindle Assembly Checkpoint Signaling Dynamics. Current Biology. Т. 25, № 20. с. R1002—R1018. doi:10.1016/j.cub.2015.08.051. Архів оригіналу за 28 червня 2018. Процитовано 25 лютого 2017.
- ↑ а б D'Archivio, Simon; Wickstead, Bill (February 2017). Trypanosome outer kinetochore proteins suggest conservation of chromosome segregation machinery across eukaryotes. The Journal of Cell Biology. Т. 216, № 2. с. 379—391. doi:10.1083/jcb.201608043. ISSN 1540-8140. PMC 5294786. PMID 28034897. Архів оригіналу за 23 липня 2018. Процитовано 19 березня 2018.
{{cite news}}: Cite має пустий невідомий параметр:|16=(довідка)Обслуговування CS1: Сторінки з PMC з іншим форматом (посилання) - ↑ а б Musacchio, Andrea (19 жовтня 2015). The Molecular Biology of Spindle Assembly Checkpoint Signaling Dynamics. Current biology: CB. Т. 25, № 20. с. R1002—1018. doi:10.1016/j.cub.2015.08.051. ISSN 1879-0445. PMID 26485365. Архів оригіналу за 26 березня 2017. Процитовано 15 березня 2017.
- ↑ Lara-Gonzalez, Pablo; Westhorpe, Frederick G.; Taylor, Stephen S. (20 листопада 2012). The spindle assembly checkpoint. Current biology: CB. Т. 22, № 22. с. R966—980. doi:10.1016/j.cub.2012.10.006. ISSN 1879-0445. PMID 23174302. Архів оригіналу за 26 березня 2017. Процитовано 15 березня 2017.
- ↑ Schleicher W. (1879). Die Knorpelzelltheilung. Ein Beitrag zur Lehre der Theilung von Gewebezellen. Archiv für Mikroskopische Anatomie. Springer-Verlag. 16 (1): 248—300. doi:10.1007/BF0295638. Архів оригіналу за 10 вересня 2014.
- ↑ Peremeschko, P.I. (1878). Über die Teilung der Zelle. Vorläufige Mitteilung. Zbl. med. Wissensch. 16 (29): 547—548.
- ↑ В. П. Шипулін, В. Д. Догузов. 160 років Анатомічному театру Університету св. Володимира. Scientific Journal of the Ministry of Health of Ukraine № 2 (3) 2013.
Джерела
ред.- А. В. Сиволоб, К. С. Афанасьєва (2012). Молекулярна організація хромосом (PDF) (укр.). К: Видавничо-поліграфічний центр «Київський університет». с. 249—293. Архів оригіналу (PDF) за 23 вересня 2015. Процитовано 25 лютого 2017.
- Bruce Alberts, Alexander Johnson, Julian Lewis, David Morgan, Martin Raff Keith Roberts, and Peter Walter (2014). Molecular biology of the cell (англ.) (вид. 6). Garland Science. с. 978—995. ISBN 9780815344322. OCLC 902708011.
- А. В. Сиволоб, С.Р. Рушковський, С.С. Кир'яченко та ін. (2008). Генетика (PDF). К: Видавничо-поліграфічний центр «Київський університет». с. 20, 27-28, 47-49. Архів оригіналу (PDF) за 4 березня 2016. Процитовано 19 березня 2016.
{{cite book}}: Явне використання «та ін.» у:|author=(довідка)
Посилання
ред.- Гарна література для зацікавлених, без складних подробиць
- Dennis,, Bray,; Karen,, Hopkin,; D.),, Johnson, Alexander (Alexander; 1946-2014,, Lewis, Julian,; C.,, Raff, Martin; (Keith),, Roberts, K.; (Professor),, Walter, Peter. Essential cell biology. ISBN 9780815344544. OCLC 893458785.
- Інше
- A Flash animation comparing Mitosis and Meiosis [Архівовано 8 березня 2021 у Wayback Machine.]
- Khan Academy, lecture
- Studying Mitosis in Cultured Mammalian Cells [Архівовано 16 травня 2008 у Wayback Machine.]
- General K-12 classroom resources for Mitosis [Архівовано 16 березня 2012 у Wayback Machine.]