Зорова кора
Зорова кора — частина кори головного мозку, яка відіграє важливу роль в обробці візуальної інформації. Вона розташована в потиличній частці, в задній частині черепа.
| Зорова кора | |
|---|---|
 Головний мозок людини. Вигляд ззаду. Червоним позначено Поле Бродмана 17 (первинну зорову кору), помаранчевим - Поле Бродмана 18 і жовтим - 'Поле Бродмана 19 | |
| Деталі | |
| Ідентифікатори | |
| Латина | Cortex visualis |
| MeSH | D014793 |
| NeuroLex ID | nlx_143552 |
| FMA | 242644 |
| Анатомічна термінологія | |
Візуальна інформація, яка надходить від очей, проходить через бічні колінчасті ядра таламусу, а потім досягає зорової кори. Частина зорової кори, яка отримує сенсорні входи від таламуса називається первинною зоровою корою, або першою візуальною зоною (V1) (відповідає Полю Бродмана 17), і екстрастріарної кори, яка складається з зорових областей другої (V2), третьої (V3), четвертої (V4), і п'ятої (V5)[1] й відповідає полям Бродмана 18 і 19.
Обидві півкулі мозку містять зорову кору. Зорова кора лівої півкулі отримує сигнали від правого зорового поля, а зорова кора в правій півкулі отримує сигнал від лівого поля зору.
Введення
ред.Первинна зорова кора (V1) знаходиться в шпорній борозні та і навколо неї, в потиличній частці. В кожній півкулі V1 отримує інформацію безпосередньо від іпсілатерального латерального колінчатого тіла , яке, в свою чергу, отримує сигнали від контрлатеральної половини поля зору.
Нейрони в зоровій корі генерують потенціали дії, коли візуальні стимули з'являються в їхньому рецептивному полі. За визначенням, рецептивне поле — це ділянка в межах поля зору, що викликає потенціал дії. Прийняті моделі нейронних налаштувань дозволяють припустити, що нейрони реагують відповідно до різних ступенів схожості між оптимальним подразником нейрона і даним стимулом. Це властивість називається нейрональним т'юнінгом. В ранніх зорових зонах нейрони мають простіші налаштування. Наприклад, нейрон в V1 може спрацювати на будь-який вертикальний стимул у його рецептивному полі. У вищих зорових зонах нейрони мають складніші налаштування. Наприклад, в нижній скроневої корі нейрон може спрацювати лише тоді, коли певне обличчя з'являється в його рецептивному полі.
Зорова кора отримує кровопостачання, насамперед, від шпорної гілки задньої мозкової артерії.
-
 На малюнку: Right Visual Field - праве зорове поле; Left Visual Field - ліве зорове поле; Right Eye - праве око; Left Eye - ліве око; Optic Nerve - оптичний нерв; Optic Chiasm - зорова хіазма; Lateral Geniculate Nucleus - бічне колінчасте ядро; Primary Visual Cortex - первинна зорова кора; Brain - мозок.
На малюнку: Right Visual Field - праве зорове поле; Left Visual Field - ліве зорове поле; Right Eye - праве око; Left Eye - ліве око; Optic Nerve - оптичний нерв; Optic Chiasm - зорова хіазма; Lateral Geniculate Nucleus - бічне колінчасте ядро; Primary Visual Cortex - первинна зорова кора; Brain - мозок. -
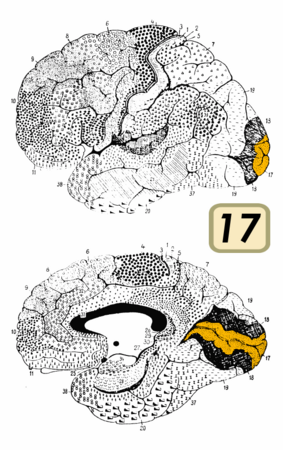 Поле Бродмана 17
Поле Бродмана 17 -
 Поле Бродмана 18
Поле Бродмана 18 -
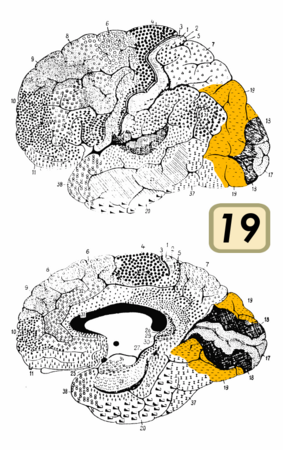 Поле Бродмана 19
Поле Бродмана 19




Сучасні дослідження
ред.Дослідження первинної зорової кори може включати в себе запис потенціалів дії від електродів у мозку котів, тхорів, щурів, мишей або мавп, або через запис вбудованих оптичних сигналів від тварин або ЕЕГ, МЕГ, або фМРТ сигнали від поля V1 людини чи мавпи .
Одним з недавніх відкриттів у відношенні зони V1 людини є те, що сигнали, виміряні за допомогою фМРТ показують дуже велику «увагову модуляцію» — зміну сигналу відповідно до уваги, сконцентрованої на подразнику. Цей результат узгоджується з іншим недавнім електрофізіологічним дослідженням, яке показало, що, хоча V1 обробляє візуальну інформацію перед V2 і V4, увагова модуляція відбувається в V4 далі — V2 і потім, нарешті, в V1. Це може означати, що увагова модуляція відбувається за рахунок зворотного зв'язку від вищих зорових областей до нижчого рівня зорових областей[2] Інші поточні дослідження зони V1 мають на меті повністю схарактеризувати його властивості підлаштовування, щоби використовувати цю ділянку як модель «канонічного» кортикального ланцюжка.
Пошкодження первинної зорової кори, як правило, призводять до скотоми, або «дірки» полі зору. Цікаво, що у пацієнти зі скотомами нерідко в змозі використовувати візуальноу інформацію, представлену в їхніх скотомах, але в змозі свідомо сприймати його. Це явище було частково описане в аспекті того, що Ларі Вайскранц (Larry Weiskrantz) з Оксфордського університету назвав «сліпим зором», і широко вивчається іншими вченими в дослідженнях свідомості.
Психологічна модель нейронної обробки візуальної інформації
ред.Вентрально-дорсальна модель
ред.
- Вентральний потік починається з V1, йде через зорові зони V2, потім через зорові зони V4 і далі, до нижньої скроневої кори. Його ще іноді називають «Що-шлях», він пов'язаний з розпізнаванням форми та визнанням об'єктів. Він також пов'язаний зі сховищем довготривалої пам'яті.
- Дорзальний потік починається з V1, йде через зорову зону V2, потім прямує до дорзомедіальної зони V6 і середньоскроневої зони V5 і далі до задньотім'яної кори. Дорзальний потік, «Де-шлях» або «Як-шлях», пов'язаний з рухом, місцезнаходженням об'єкта, контролем ока і руки, особливо, коли візуальна інформація використовується для орієнтації саккад (узгоджених рухів очей для розпізнавання об'єкту, обличчя людини) чи досягнення об'єкту рукою.
«Що-» і «де-» функції вентрального й дорзального шляхів були вперше описані Унгерлайдером і Мішкіним (Ungerleider and Mishkin)[3]
Ще зовсім недавно, Гудейл (Goodale) і Мілнер (Milner) висловили ці ідеї й припустили, що вентральний потік має вирішальне значення для візуального сприйняття, тоді як дорсальний потік опосередковує візуальний контроль відпрацьованих дій.[4]Було показано, що зорові ілюзії, такі як ілюзія Еббінгауза (англ. Ebbinghaus illusion) спотворюють уявлення перцептивної природи, але, коли випробуваний відповідає дією, наприклад, хапанням, ніякого спотворення не відбувається.[5]
В одному з досліджень (Scharnowski і Gegenfurtner)[6] припускається, що і система дії, й система сприйняття однаково піддаються таким ілюзіям. Інші дослідження, тим не менш, більше підтримують ідею, що на такі відпрацьовані дії, такі як хапання не впливають ілюзії[7][8]і зазначають, що дисоціації дії/сприйняття є корисним прикладом, для описання функціонального розподілу праці між дорсальним і вентральним зоровими шляхами в корі головного мозку.[9]
Первинна зорова кора (V1)
ред.

Первинна зорова кора — найкраще вивчена зорова зона в мозку У всіх досліджених ссавців, вона знаходиться у задньому полюсі потиличної частки. Це найпростіша, найбільш рання кіркова зорова зона, спеціалізована для обробки інформації про статичні й рухомі об'єкти, й для розпізнавання самих об'єктів
Функціонально визначена зона первинної зорової кори приблизно відповідає анатомічно визначеній смугастій (стріарній) корі. Назва « смугаста кора» походить від лінії Геннарі, смуги, яка видна неозброєним оком[10] що являє собою мієлінізовані аксони з бічного колінчатого ядра, які закінчуються в шарі IV сірої речовини.
Первинна зорова кора складається із шести функціонально різних шарів, пронумерованих від 1 до 6. Середнє число нейронів, які містяться в первинній зоровій корі дорослої людини, у кожній півкулі, оцінюється в розмірі близько 140 мільйонів.[11]
Функція
ред.V1 має дуже чітко визначені карти просторової інформації. Наприклад, у людини, верхній край шпорної борозни сильно реагує на інформацію з нижньої половині поля зору (нижче центру), а нижній край шпорної борозни — на інформацію з верхньої половини поля зору. Так відбувається трансформація візуального образу від сітківки до V1. Відповідність між цим об'єктом у V1 і в суб'єктивному полі зору дуже точне: навіть сліпі плями відображаються в V1. З точки зору еволюції, ця передача зображення — базова і є у більшості тварин, які мають V1. У людини і тварин з центральною ямкою в сітківці, більша частина V1 відображає невелику центральну ділянку поля зору. Це явище знане як кортикальне збільшення (англ. cortical magnification) .[12] Можливо, з метою точного просторового відображення, нейрони в V1 мають найменший розмір рецептивного поля з усіх ділянок зорової кори.
Властивості V1 нейронів та їхнє реагування значно відрізнятися з плином часу. На початку часу (40 ms і далі) окремі нейрони V1 добре підлаштовані до невеликого набору подразників. Тобто, нейронні реакції можуть розрізняти невеликі зміни візуальної орієнтації, просторової частоти і кольору. Крім того, окремі V1 нейрони людини й тварин з бінокулярним зором мають монокулярне домінування, тобто, налаштування на одне з двох очей. У V1 і первинної сенсорної кори в цілому, нейрони з таким характерним налаштуванням мають тенденцію групуватися разом, в кортикальні колонки. Девід Хьюбел і Торстен Візел запропонували класичну модель організації кортикальної колонки у вигляді «кубику льоду» для пояснення двох властивостей такого налаштування: монокулярного домінування[en] й орієнтації[en]. Однак ця модель не може вмістити кольору, просторової частоти і багатьох інших функцій, до яких налаштовані нейрони . Точна модель організації всіх цих кортикальних колонок в V1 залишається актуальною темою сучасних досліджень. Математичне моделювання цієї функції було зроблено на основі моделі перетворення Гарбора (англ. Gabor transforms).
Візуальна інформація ретранслюється на V1 не закодовано у термінах просторових (або оптичних) зображень, а, швидше, у вигляді контурів. Як приклад, для зображень, що скомпоновані з чорної й білої половин, розділова межа між чорним та білим являє собою сильний локальний контраст і кодується, в той самий час, поки декілька нейронів кодують інформацію про яскравість (як чорного, так і білого кольору). Інформація додатково передаються на наступні зорові ділянки.[13] Функціональні моделі простих клітин в V1 було встановлено Ліндебергом (Lindeberg) та іншими вченими[14][15][16][17][18]
Розбіжності в розмірах V1 також впливають на індивідуальне сприйняття й ілюзії[19]
V2
ред.Зорова зона V2, або вторинна зорова кора, яка ще називається престріарною або предсмугастою корою -[20] друга за значенням ділянкою в зоровій корі, і першою ділянкою в полі візуальних асоціацій. Вона має потужні прямі зв'язки з V1 (прямі й через подушку зорового бугра) і спрямовує волокна до V3, V4 і V5. Зона також має міцний зворотний зв'язок з V1.
З точки зору анатомії, V2 поділяється на чотири квадранти — дорсальне і вентральне представництво в лівій і правій півкулі. Разом, ці чотири регіони забезпечують повну карту візуального світу. V2 має багато спільних з V1 властивостей: клітини налаштовані на прості властивості, такі як орієнтація, просторова частота і колір. Відповіді на багатьох нейронів V2 також модулюються складнішими феноменами, такими як ілюзорні контури,[21][22] бінокулярна невідповідність,[23] в залежності від того стимул є частиною малюнка чи частиною тла.[24][25] недавні дослідження показали, що для клітин зони V2 вплив уваги (увагова модуляція) невеликий. Він більший ніж в зоні V1, але менший, ніж у зоні V4. При цьому клітини налаштовані на помірно складні патерни й можуть бути керовані множинними установками в різних субрегіонах одного рецептивного поля. Дослідники стверджують, що весь вентральний зорово-гіпокампальний потік важливий для зорової пам'яті.[26]ця теорія, на відміну від домінуючої, стверджує, що зміни об'єктно-розпізнавальної пам'яті (ОРП) можуть стати результатом внутрішніх процесів у зоні V2, яка має тісні взаємозв'язки в межах вентрального потоку зорової кори. В мозку мавпи, ця ділянка має сильні прямі з'єднання з первинною зоровою корою (V1) і посилає імпульси на інші вторинні ділянки зорової кори (V3, V4 і V5).[27][28][22][29][30]
У недавньому дослідженні, шар 6 клітин кори V2, як виявилося, грає дуже важливу роль у зберіганні розпізнаного об'єкту пам'яті, а також перетворення об'єкту короткострокової спогадів в об'єкт довгострокової пам'яті.[31]
Третя зорова кора, включно з V3
ред.Термін третій візуальний комплекс стосується області кори, розташованої безпосередньо перед V2, і який включає в області візуальної зони V3 в мозку людини. «Ускладнена» номенклатура обумовлена тим, що деякі розбіжності відносно точних розмірів зони V3 все ще існують деякі дослідники, припускаючи, що ділянка кори, розташована в передній частині V2, може включати в себе два або три функціональних підрозділи. Наприклад, Девід Ван Ессен (David Van Essen) та інші (1986) запропонували існування «дорзальної V3» у верхній частині мозкового півкулі, яка відрізняється від «вентральної V3» , розташованої в нижній частині мозку. Дорзальна й вентральна V3 мають чіткі зв'язки з іншими частинами мозку, з'являються в різних зрізах, забарвлених різними барвниками за різними методиками й містять нейрони, які реагують на різні комбінації візуальних стимулів (наприклад, кольорово-селективні нейрони, більш поширені у вентральній V3). В мозку людини були виділені також додаткові підрозділи, в тому числі V3A і V3B. Ці підрозділи знаходяться поруч з вентральною V3, але не прилягають до V2. Дорзальна V3, яка зазвичай розглядається як частина дорзального потоку, отримує входи від V2 і від первинної зорової кори й сама проєктує інформацію до задньої тім'яної кори. Це може бути анатомічно розташовані в полі Бродмана 19. Braddick, використовуючи фМРТ припустив, що зона V3/V3A може грати роль у загальному сприйнятті руху[32] інші дослідження воліють розглядати дорзальну V3 як частину більш великої дорзомедіальної зони (ДМ), який містить представництво всього поля зору. Нейрони в області ДМ реагують на когерентний рух великих об'єктів, що охоплюють значні частини поля зору (Lui і співавтори, 2006).
Вентральна V3, має набагато слабші з'єднання з первинною зоровою зоною, і тісніші зв'язки з нижньою скроневою корою. У той час як у більш ранніх дослідженнях стверджувалося, що у V3 міститься інформація тільки з верхньої частини поля зору (вище точки фіксації), пізніші роботи показали, що ця область ширша і, як в інших зорових зонах, у ній може міститися повне візуальне подання[33]
V4
ред.Зорова зона V4 — одна із зорових зон в екстрастріарній зоровій корі. У макак, вона розташована спереду від V2 і ззаду від задньотім'яної ділянки. Зона включає як мінімум чотири ділянки (ліва й права V4d, ліва й права V4v), а деякі дослідники зазначають, що V4 містить також ростральні і каудальні підрозділи. Невідомо достеменно, чи існує гомологічна до V4 ділянка в мозку людини, і це питання на даний час — предмет пильної уваги.[34]
Більшість досліджень вказують на те, що вибіркова увага може змінити показники імпульсації в V4 приблизно на 20 %..[35][36]
Середньотім'яна зорова зона (V5)
ред.Середньотім'яна зорова зона (МТ або V5)- це ділянка екстрастріарної зорової кори. У декількох видів широконосих мавп і мавп Старого Світу МТ зона містить високу кількість напрямок-селективних нейронів.[37] МТ приматів відіграє основну роль у сприйнятті руху, інтеграції локальних рухових сигналів у глобальне сприйняття, й керуванні рухом очей[37]
Зв'язки
ред.МТ під'єднана до широкого спектра кортикальних і підкіркових областей головного мозку. Імпульси до цієї зони надходять із візуальних областей кори головного мозку V1, V2 і V3 (дорсомедиальнї зони),[38][39] кіноцелюлярних зон,[40] і нижньої подушки зорового бугра.[41] Патерн проєкції на МТ робить зміни в сприйнятті центрального і периферичного поля зору.[42]
Стандартним є уявлення про те, що V1 є «найбільш важливим» входом у MT[37] Однак, кілька досліджень показали, що нейрони в МТ здатні реагувати на візуальну інформацію, часто в напрям-селективному режимі, навіть після знищення або інактивації V1.[43] крім того, дослідження Семіра Зекі (Semir Zeki) й співавторів показали, що певні типи візуальної інформації можуть досягати МТ ще до того, як потраплять до V1.
МТ відправляє свої основні імпульси до ділянок розташованих в корі головного мозку, які безпосередньо оточують його, в тому числі в районах FST, MST, і V4t . Інші виходи з МТ прямують до зон руху очей в лобовій та тім'яній частках.
Функція
ред.Перші дослідження електрофізіологічних властивостей нейронів в зоні МТ показали, що велика частина її клітин налаштовані на швидкість і напрям руху зорових стимулів.[44][45]
Дослідження з пошкодженням також підтвердили роль МТ в сприйнятті рухів і рухах очей.[46] Нейропсихологічні дослідження пацієнта, який не в змозі побачити рух, який дивиться на світ як на серію статичних кадрів, привели до думки, що зона V5 у приматів гомологічна МТ у людини.[47][48]
Однак, оскільки нейрони в V1, також налаштовані на напрям і швидкість руху, ці ранні результати залишають відкритим питання про те, що саме МТ може такого зробити, що не зможе зробити V1. Вчені проробили великий обсяг роботи в дослідженні цієї ділянки, аж допоки стало зрозуміло, що саме тут відбувається інтеграція локальних візуальних сигналів руху в глобальні рухи складних об'єктів.[49] Наприклад, пошкодження V5 веде до появи дефіциту в сприйнятті руху і обробки складних подразників. V5 містить багато нейронів, які вибірково обробляють рух складних візуальних об'єктів (кінці ліній, кутів). Мікрополяризація нейрона, розташованого в V5 впливає на сприйняття руху.[50]
Є ще багато суперечок з приводу точної форми обробки інформації в зоні МТ[51] і деякі дослідження показують, що сприйняття руху, по суті, вже існує на нижчих рівнях зорової системи, таких як V1. [52] [53]
Функціональна організація
ред.Було доведено, що нейрони в МТ організовані в правильні стовпчики.[54] DeAngelis стверджував, що в МТ нейрони також організовані на основі їх бінокулярної диспаратності (невідповідності).[55]
V6
ред.Дорсомедиальна зона (ДМ), також знана як зона V6, вбачається, що реагує на візуальні стимули, пов'язані з власним рухом[56] і ширококутніми (в полі зору) об'єктами.[57] V6, як структурний підрозділ зорової кори приматів, уперше описав Джон Оллман і Джон Каас (John Allman and Jon Kaas) у 1975 році.[58]V6 знаходиться у дорзальній частині екстрастріарної кори, поруч з поздовжньою щілиною великого мозку (лат. fissura longitudinalis medialis), і, як правило, також включає в себе частини медіальної кори, такі, як тім'яно-потилична борозна.
Протягом багатьох років вважалося, що ДМ існує тільки у широконосих мавп (мавп Нового Світу). Однак пізніші дослідження показали, що ДМ існує і в мавп Старого Світу і, можливо, у людей. V6 також іноді називають тім'яно-потиличною зоною , хоча відповідність не точна.[59][60]
Властивості
ред.Нейрони в зоні ДМ/V6 нічної мавпи і ігрунки звичайної мають унікальні властивості реагування, такі як надзвичайно висока вибірковість до орієнтації візуальних контурів, і перевагу довгих, безперервних ліній, що охоплюють значні частини поля зору[61][62]
Проте, в порівнянні з зоною MT, значно менша частка клітин ДМ показує вибіркову відповідь на напрямок руху візуальних образів. Ще однією помітною різницею з МТ є те, що клітини в ДМ налаштовані на низькі просторові частоти зображення й погано реагують на рухи фактурних об'єктів, таких як поле випадкових точок. Ці реакції припускають, що ДМ і МТ можуть працювати паралельно: перша — аналізуючи власний рух по відношенню до навколишнього середовища, а друга — аналізуючи рухи окремих об'єктів щодо тла.
Нещодавно зона, яка реагує на ширококутні зорові поля, була визначена у людини, і вважається, що вона гомологічна зоні V6 макаки.[63]
Шляхи
ред.Зв'язки та властивості реагування клітин в DM / V6 дозволяють припустити, що ця область є ключовою в складі дорзального потоку, названого деякими дослідниками «дорсомедиальним шляхом». Цей шлях, ймовірно, важливий для контролю скелетомоторної активності, в тому числі постуральних реакцій і рухів, спрямованих на «досягання об'єктів»[60] Ця зона має відносно прямі зв'язки з регіонами лобової частки, що контролюють рухи руки, в тому числі і з премоторною корою (частиною поля Бродмана 6).
Див. також
ред.Примітки
ред.- ↑ Mather, George. The Visual Cortex. School of Life Sciences: University of Sussex (English) . University of Sussex. Архів оригіналу за 3 лютого 2017. Процитовано 6 березня 2017.
- ↑ Buffalo, E.A., Fries, P., Landman, R., Liang, H., and Desimone, R. (2010). A backward progression of attentional effects in the ventral stream. Proceedings for the National Academy of Sciences. 107(1), 361—365.
- ↑ Ungerleider LG, Mishkin M (1982). Two Cortical Visual Systems. У Ingle DJ, Goodale MA, Mansfield RJ (ред.). Analysis of Visual Behavior. Boston: MIT Press. с. 549—586.
- ↑ Goodale MA, Milner AD (1992). Separate pathways for perception and action. Trends in Neuroscience. 15 (1): 20—25. doi:10.1016/0166-2236(92)90344-8. PMID 1374953.
- ↑ Aglioti S, DeSouza JF, Goodale MA (1995). Size-contrast illusions deceive the eye but not the hand. Curr. Biol. 5 (6): 679—85. doi:10.1016/S0960-9822(95)00133-3. PMID 7552179.
- ↑ Franz VH, Scharnowski F, Gegenfurtner (2005). Illusion effects on grasping are temporally constant not dynamic. J Exp Psychol Hum Percept Perform. 31 (6): 1359—78. doi:10.1037/0096-1523.31.6.1359. PMID 16366795.
- ↑ Ganel T, Goodale MA (2003). Visual control of action but not perception requires analytical processing of object shape. Nature. 426 (6967): 664—7. doi:10.1038/nature02156. PMID 14668865.
- ↑ Ganel T, Tanzer M, Goodale MA (2008). A double dissociation between action and perception in the context of visual illusions: opposite effects of real and illusory size. Psych. Sci. 19 (3): 221—5. doi:10.1111/j.1467-9280.2008.02071.x. PMID 18315792.
- ↑ Goodale MA. (2011). Transforming vision into action. Vision Res. 51 (14): 1567—87. doi:10.1016/j.visres.2010.07.027. PMID 20691202.
- ↑ Glickstein M., Rizzolatti G. Francesco Gennari and the structure of the cerebral cortex Trends in Neurosciences, Volume 7, Issue 12, 464—467, 1 December 1984.
- ↑ Leuba G; Kraftsik R (1994). Changes in volume, surface estimate, three-dimensional shape and total number of neurons of the human primary visual cortex from midgestation until old age. Anatomy and Embryology. 190 (4): 351—366. doi:10.1007/BF00187293. PMID 7840422.
- ↑ Barghout, Lauren (1999). On the Differences Between Peripheral and Foveal Pattern Masking. Berkeley, California, U.S.A.: Masters Thesis. U.C. Berkeley.
- ↑ Barghout, Lauren (2003). Vision: How Global Perceptual Context Changes Local Contrast Processing (Ph.D. Dissertation). Updated to include computer vision techniques. Scholar's Press. ISBN 978-3-639-70962-9. Архів оригіналу за 18 листопада 2016. Процитовано 17 березня 2017.
- ↑ Lindeberg, T. A computational theory of visual receptive fields. Biological Cybernetics. 107 (6): 589—635. doi:10.1007/s00422-013-0569-z.
- ↑ Lindeberg, T. Time-causal and time-recursive spatio-temporal receptive fields. Journal of Mathematical Imaging and Vision. 55 (1): 50—88. doi:10.1007/s10851-015-0613-9. Архів оригіналу за 18 березня 2017. Процитовано 17 березня 2017.
- ↑ DeAngelis, G. C.; Ohzawa, I.; Freeman, R. D. (1995). Receptive field dynamics in the central visual pathways. Trends Neurosci. 18 (10): 451—457. doi:10.1016/0166-2236(95)94496-r. PMID 8545912.
- ↑ G. C. DeAngelis and A. Anzai "A modern view of the classical receptive field: linear and non-linear spatio-temporal processing by V1 neurons. In: Chalupa, L.M., Werner, J.S. (eds.) The Visual Neurosciences, vol. 1, pp. 704—719. MIT Press, Cambridge, 2004.
- ↑ Lindeberg, T. Invariance of visual operations at the level of receptive fields. PLOS ONE. 8 (7): e66990. doi:10.1371/journal.pone.0066990.
{{cite journal}}: Обслуговування CS1: Сторінки із непозначеним DOI з безкоштовним доступом (посилання) - ↑ Schwarzkopf, D Samuel (2011). The surface area of human V1 predicts the subjective experience of object size. Nature Neuroscience. 14 (1): 28—30. doi:10.1038/nn.2706. PMC 3012031. PMID 21131954. Архів оригіналу за 1 вересня 2011. Процитовано 17 березня 2017.
- ↑ Gazzaniga, Ivry & Mangun: Cognitive neuroscience, 2002
- ↑ von der Heydt, R; Peterhans, E; Baumgartner, G (1984). Illusory contours and cortical neuron responses. Science. 224: 1260—62. doi:10.1126/science.6539501. PMID 6539501.
- ↑ а б Anzai, A; Peng, X; Van Essen, D. C (2007). Neurons in monkey visual area V2 encode combinations of orientations. Nature Neuroscience. 10 (10): 1313—21. doi:10.1038/nn1975. PMID 17873872.
- ↑ von der Heydt, R; Zhou, H; Friedman, H. S (2000). Representation of stereoscopic edges in monkey visual cortex. Vision Research. 40: 1955—67. doi:10.1016/s0042-6989(00)00044-4.
- ↑ Qiu, F. T; von der Heydt, R (2005). Figure and ground in the visual cortex: V2 combines stereoscopic cues with Gestalt rules. Neuron. 47: 155—66. doi:10.1016/j.neuron.2005.05.028.
- ↑ Maruko, I; et alt. (2008). Postnatal Development of Disparity Sensitivity in Visual Area 2 (V2) of Macaque Monkeys. Journal of Neurophysiology. 100 (5): 2486—2495. doi:10.1152/jn.90397.2008.
- ↑ Bussey, T J; Saksida, L. M (2007). Memory, perception, and the ventral visual-perirhinal-hippocampal stream: thinking outside of the boxes. Hippocampus. 17 (9): 898—908. doi:10.1002/hipo.20320. PMID 17636546.
- ↑ Stepniewska, I; Kaas, J. H. (1996). Topographic patterns of V2 cortical connections in macaque monkeys. The Journal of Comparative Neurology. 371 (1): 129—152. doi:10.1002/(SICI)1096-9861(19960715)371:1<129::AID-CNE8>3.0.CO;2-5. PMID 8835723.
- ↑ Gattas, R; Sousa, A. P; Mishkin, M; Ungerleider, L. G. (1997). Cortical projections of area V2 in the macaque. Cerebral Cortex. 7 (2): 110—129. doi:10.1093/cercor/7.2.110.
- ↑ Hegdé, Jay; Van Essen, D. C (2000). Selectivity for Complex Shapes in Primate Visual Area V2. The Journal of Neuroscience. 20. Архів оригіналу за 24 жовтня 2016. Процитовано 17 березня 2017.
- ↑ Hegdé, Jay; Van Essen, D. C (2004). Temporal dynamics of shape analysis in Macaque visual area V2. Journal of Psychology. 92 (5): 3030—3042. doi:10.1152/jn.00822.2003.
- ↑ López-Aranda et alt. (2009). Role of Layer 6 of V2 Visual Cortex in Object Recognition Memory. Science. 325 (5936): 87—89. doi:10.1126/science.1170869. PMID 19574389.
- ↑ Braddick, OJ, O'Brien, JM та ін. (2001). Brain areas sensitive to coherent visual motion. Perception. 30 (1): 61—72. doi:10.1068/p3048. PMID 11257978.
- ↑ Rosa, MG; Tweedale, R (2000). Visual areas in lateral and ventral extrastriate cortices of the marmoset monkey. J Comp Neurol. 422 (4): 621—51. doi:10.1002/1096-9861(20000710)422:4<621::AID-CNE10>3.0.CO;2-E. PMID 10861530.
- ↑ Goddard E; Goddard, Erin; McDonald, J. S.; Solomon, S. G.; Clifford, C. W. G. (2011). Color responsiveness argues against a dorsal component of human V4. Journal of Vision. 11 (4): 3. doi:10.1167/11.4.3. PMID 21467155.
- ↑ http://www.sciencemag.org/cgi/content/abstract/229/4715/782)
- ↑ Moran, J; Desimone, R (1985). Selective Attention Gates Visual Processing in the Extrastriate Cortex. Science. 229 (4715): 782—4. doi:10.1126/science.4023713. PMID 4023713.
- ↑ а б в Born R, Bradley D (2005). Structure and function of visual area MT. Annu Rev Neurosci. 28: 157—89. doi:10.1146/annurev.neuro.26.041002.131052. PMID 16022593.
- ↑ Felleman D, Van Essen D (1991). Distributed hierarchical processing in the primate cerebral cortex. Cereb Cortex. 1 (1): 1—47. doi:10.1093/cercor/1.1.1-a. PMID 1822724.
- ↑ Ungerleider L, Desimone R (1986). Cortical connections of visual area MT in the macaque. J Comp Neurol. 248 (2): 190—222. doi:10.1002/cne.902480204. PMID 3722458.
- ↑ Sincich L, Park K, Wohlgemuth M, Horton J (2004). Bypassing V1: a direct geniculate input to area MT. Nat Neurosci. 7 (10): 1123—8. doi:10.1038/nn1318. PMID 15378066.
- ↑ Warner CE, Goldshmit Y, Bourne JA (2010). Retinal afferents synapse with relay cells targeting the middle temporal area in the pulvinar and lateral geniculate nuclei. Front Neuroanat. 4: 8. doi:10.3389/neuro.05.008.2010. PMC 2826187. PMID 20179789.
{{cite journal}}: Обслуговування CS1: Сторінки із непозначеним DOI з безкоштовним доступом (посилання) - ↑ Palmer SM, Rosa MG (2006). A distinct anatomical network of cortical areas for analysis of motion in far peripheral vision. Eur J Neurosci. 24 (8): 2389—405. doi:10.1111/j.1460-9568.2006.05113.x. PMID 17042793.
- ↑ Rodman, HR; Gross, CG; Albright, TD (1989). Afferent basis of visual response properties in area MT of the macaque. I. Effects of striate cortex removal. J Neurosci. 9 (6): 2033—50. PMID 2723765.
- ↑ Dubner R, Zeki S (1971). Response properties and receptive fields of cells in an anatomically defined region of the superior temporal sulcus in the monkey. Brain Res. 35 (2): 528—32. doi:10.1016/0006-8993(71)90494-X. PMID 5002708..
- ↑ Maunsell J, Van Essen D (1983). Functional properties of neurons in middle temporal visual area of the macaque monkey. I. Selectivity for stimulus direction, speed, and orientation. J Neurophysiol. 49 (5): 1127—47. PMID 6864242.
- ↑ Dursteler M.R.; Wurtz R.H.; Newsome W.T. (1987). Directional pursuit deficits following lesions of the foveal representation within the superior temporal sulcus of the macaque monkey. Journal of Neurophysiology. 57 (5): 1262—87. PMID 3585468.
- ↑ Hess R.H.; Baker C.L.; Zihl J. (1989). The 'motion-blind' patient: low-level spatial and temporal filters. Journal of Neuroscience. 9 (5): 1628—40. PMID 2723744.
- ↑ Baker C.L. Jr; Hess R.F; Zihl J. (1991). Residual motion perception in a 'motion-blind' patient, assessed with limited-lifetime random dot stimuli. Journal of Neuroscience. 11 (2): 454—61. PMID 1992012.
- ↑ Movshon, J.A., Adelson, E.H., Gizzi, M.S., & Newsome, W.T. (1985). The analysis of moving visual patterns. In: C. Chagas, R. Gattass, & C. Gross (Eds.), Pattern recognition mechanisms (pp. 117—151), Rome: Vatican Press.
- ↑ Britten K.H.; van Wezel R.J. (1998). Electrical microstimulation of cortical area MST biases heading perception in monkeys. Nat Neurosci. 1 (1): 59—63. doi:10.1038/259. PMID 10195110.
- ↑ Wilson, H.R.; Ferrera, V.P.; Yo, C. (1992). A psychophysically motivated model for two-dimensional motion perception. Vis Neurosci. 9 (1): 79—97. doi:10.1017/s0952523800006386.
- ↑ Tinsley, C.J., Webb, B.S., Barraclough, N.E., Vincent, C.J., Parker, A., & Derrington, A.M. (2003). The nature of V1 neural responses to 2D moving patterns depends on receptive-field structure in the marmoset monkey. J Neurophysiol. 90 (2): 930—7. doi:10.1152/jn.00708.2002. PMID 12711710.
- ↑ Pack C.C.; Born R.T.; Livingstone M.S. (2003). Two-dimensional substructure of stereo and motion interactions in macaque visual cortex. Neuron. 37 (3): 525—35. doi:10.1016/s0896-6273(02)01187-x. PMID 12575958.
- ↑ Albright T (1984). Direction and orientation selectivity of neurons in visual area MT of the macaque. J Neurophysiol. 52 (6): 1106—30. PMID 6520628.
- ↑ DeAngelis G, Newsome W (1999). Organization of disparity-selective neurons in macaque area MT. J Neurosci. 19 (4): 1398—415. PMID 9952417.
- ↑ Cardin, V; Smith, AT (2010). Sensitivity of human visual and vestibular cortical regions to stereoscopic depth gradients associated with self-motion. Cerebral Cortex. 20 (8): 1964—73. doi:10.1093/cercor/bhp268.
- ↑ Pitzalis et alt. (2006). Wide-Field Retinotopy Defines Human Cortical Visual Area V6. The Journal of Neuroscience. 26 (30): 7962—73. doi:10.1523/jneurosci.0178-06.2006. PMID 16870741.
- ↑ Allman JM, Kaas JH (1975). The dorsomedial cortical visual area: a third tier area in the occipital lobe of the owl monkey (Aotus trivirgatus). Brain Res. 100 (3): 473—487. doi:10.1016/0006-8993(75)90153-5.
- ↑ Galletti C та ін. (2005). The relationship between V6 and PO in macaque extrastriate cortex. Eur J Neurosci. 21: 959—970. doi:10.1111/j.1460-9568.2005.03911.x.
- ↑ а б Galletti C та ін. (2003). Role of the medial parieto-occipital cortex in the control of reaching and grasping movements. Exp Brain Res. 153: 158—170. doi:10.1007/s00221-003-1589-z.
- ↑ Baker JF та ін. (1981). Visual response properties of neurons in four extrastriate visual areas of the owl monkey (Aotus trivirgatus): a quantitative comparison of medial, dorsomedial, dorsolateral, and middle temporal areas. J Neurophysiol. 45: 397—416.
- ↑ Lui LL та ін. (2006). Functional response properties of neurons in the dorsomedial visual area of New World monkeys (Callithrix jacchus). Cereb Cortex. 16 (2): 162—177. doi:10.1093/cercor/bhi094.
- ↑ Pitzalis, S., Sereno, M.I., Committeri, G., Fattori, P., Galati, G., Patria, F., & Galletti, C. (2010). Human v6: The medial motion area. Cereb Cortex. 20 (2): 411—424. doi:10.1093/cercor/bhp112.
Посилання
ред.- The Primary Visual Cortex by Matthew Schmolesky Університет штата Юта
- Architecture of the Visual Cortex, by David Hubel at Гарвардський Університет
- Симулятор моделювання візуальних карт [Архівовано 9 січня 2014 у Wayback Machine.] На topographica.org