Еукаріоти
| Еукаріоти Час існування: Орозирій — наш час 1850–0 Ma | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
 Клітина еукаріотів
| ||||||||||
| Біологічна класифікація | ||||||||||
| ||||||||||
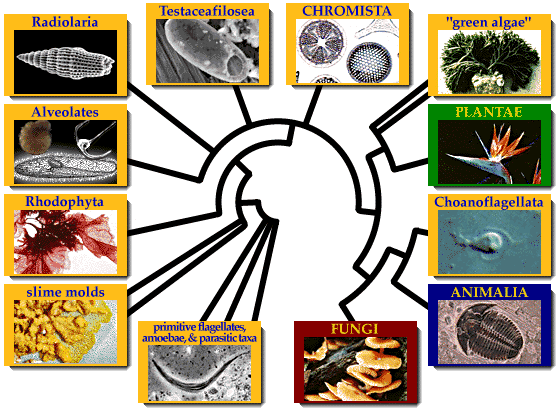
Супергрупи та царства
| ||||||||||
Посилання
| ||||||||||
|
Еукаріо́ти[1] (інколи евкаріо́ти, від грец. εύ- — повністю, добре й грец. κάρυον — ядро) або я́дерні (лат. Eukaryota Whittaker & Margulis, 1978) — домен одно- та багатоклітинних організмів, що характеризуються переважно полігеномними клітинами, морфологічно сформованим ядром та наявністю мембранних субклітинних органел.
Геноми еукаріотичної клітини представлені:
- а) ядерним геномом, зосередженим у ядрі і представленим ядерною ДНК;
- б) у більшості клітин мітохондріальним геномом, зосередженим у мітохондріях і представленим мітохондріальною ДНК;
- в) у деяких клітин пластидним (зокрема, хлоропластним) геномом, що розташовується у пластиді, і представлений хлоропластною ДНК (генофором);
- г) геномом нуклеоморфу[2], який виявлений лише у кількох відділах водоростей у надзвичайно цікавій ядроподібній структурі, розташованій між оболонками пластиди та особливою клітинною системою — хлоропластним ендоплазматичним ретикулумом. У нуклеоморфі виявлена власна, нуклеоморфна ДНК.
Двогеномні клітини, в яких є ядерний та мітохондріальний геноми, характерні для грибів та тварин; тригеномні — з ядерним, мітохондріальним та пластидним геномами — для майже всіх рослин; чотиригеномні — з ядерним, мітохондріальним, пластидним та нуклеоморфними геномами, виявлені у хлорарахніофітових та криптофітових водоростей.
Морфологія еукаріотичної клітини ред.
Клітини прокаріотів (бактерій та архей) та еукаріотів на фенотипічному рівні схожі за наявністю ДНК та білок-синтезуючого апарату, представленого рибосомами; наявністю клітинних мембран; ферментних комплексів, що забезпечують процеси реплікації, транскрипції, трансляції та синтезу АТФ. До складу клітин прокаріотів та еукаріотів входять білки, жири, вуглеводи, нуклеїнові кислоти, мінеральні речовини та вода.
Відмінності в будові клітин прокаріотів та еукаріотів пов'язані, в першу чергу, з тим, що майже вся ДНК прокаріотів складається з однієї хромосоми, і, у випадку бактерій, не пов'язана з білками-гістонами (хоча навіть бактерії мають гістоноподібні білки). Еукаріоти, на відміну від прокаріотів, зазвичай не містять генів, організованих у великі оперони, процес цитокінезу (поділу ДНК) проходить за участю веретена поділу. Рибосоми у еукаріотів більші, ніж рибосоми прокаріотів (умовна вага прокаріотичних рибосом становить 70S, тоді як еукаріотичних — 80S). Багато клітин еукаріотів здатні до фаго- та піноцитозу, мають морфологічно оформлене ядро, часто також мітохондрії, пластиди, ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, пероксисоми, мають іншу структуру джгутиків і джгутикових моторів, мають клітинний центр з центріолями. Для багатьох клітин еукаріотів характерні мітоз, мейоз, повноцінний статевий процес, проте набагато нижча ймовірність горизонтального переносу генів. Зазвичай еукаріотичні клітини більші за розмірами, ніж прокаріотичні — їхній середній діаметр становить близько 5—20 мікрон, тоді як у бактерії та архей — 0,5—2 мікрона.
Гіпотези походження еукаріотів ред.
За палеонтологічними даними, прокаріоти на нашій планеті виникли близько 3,2 млрд років тому, тоді як еукаріоти набагато молодші — їх вік становить лише близько 1,6 млрд років[3]. Сьогодні в межах доменів бактерій та архей описано близько 40 тис. видів (хоча оцінки їх загальної кількості складають близько 10 млн видів[4]), тоді як у домені еукаріотів — близько 1,7 млн видів[3].
Першим відомим евкаріотом є наземний організм Diskagma buttonii, гіпотетичний вид грибів, рештки якого було знайдено у Південній Африці.[5]
Автогенетична гіпотеза ред.
Згідно з цією гіпотезою, еукаріотична клітина утворилась з прокаріотичної внаслідок розвитку плазмалемою системи інвагінацій, які надалі замкнулись навколо ділянок плазмалеми з ферментами дихального ланцюга, фотосинтетичними пігментами, нуклеоїда, відповідно утворивши мітохондрії, пластиди та ядро. Розвиток системи інвагінацій мембран призвів також до виникнення ендоплазматичної сітки, комплексу Гольджі, лізосом та ін. Проте автогенетична гіпотеза не змогла пояснити чимало фактів. Наприклад, чому мітохондрії та хлоропласти містять власну ДНК і розмножуються поділом, чому в мітохондріях та хлоропластах наявні легкі прокаріотичні рибосоми, чому поровий апарат мітохондрій та пластид подібний до порового апарату прокаріот і відрізняється від порового апарату плазмалеми еукаріот, як виник мітоз та з'явились мікротрубочки та ін.
Ендосимбіотична гіпотеза ред.
Наприкінці XIX ст. видатний ботанік та фізіолог рослин М. С. Цвєт, на основі спостережень поділу хлоропласту та ядерця у зеленої водорості Spirogyra висловив припущення, що клітини рослин, можливо, являють собою симбіотичний утвір із клітини-господаря та бактеріальних внутрішньоклітинних симбіонтів, які відомі як хлоропласти та ядерця. Це припущення у 20—40-х роках XX ст. було детально розроблено, в першу чергу, К. С. Мережковським у так звану ендосимбіотичну гіпотезу походження еукаріотичної клітини. Проте ідеї К. С. Мережковського не знайшли підтримки у широкого загалу біологів. Гіпотезу ендосимбіотичного походження еукаріотів називали «вражаючим витвором людської фантазії», «луна-парком», «непристойною темою у колі освічених біологів»[3]. У 60-х роках гіпотеза ендосимбіотичного походження еукаріотичної клітини була заново сформульована американською дослідницею Л. Маргеліс.
Згідно з гіпотезою Мережковського—Маргеліс, еукаріотична клітина виникла внаслідок кількох ендосимбіозів: гіпотетична прокаріотична анаеробна клітина, здатна до фагоцитозу, захопила, проте не перетравила, а зберегла в цитоплазмі, аеробну гетеротрофну бактерію, яка трансформувалась у мітохондрію. Далі клітина-господар, що містила мітохондрію, вступила в симбіоз із рухливою спірохетоподібною гетеротрофною бактерією, яка дала початок джгутику. Після цього внаслідок автогенетичного процесу, пов'язаного з утворенням глибоких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря утворилось ядро. Далі базальні тіла джгутиків трансформувались в центри організації веретена поділу, внаслідок чого виник спочатку мітоз, а потім його модифікація — мейоз, разом зі статевим процесом. Виникла перша гетеротрофна еукаріотична клітина. Нащадки цієї клітини, еволюція яких пішла по шляху вдосконалення фагоцитозу, дали початок тваринам. Ті ж нащадки, які еволюціонували в напрямку удосконалення осмотрофного живлення, дали початок грибам. Симбіоз гетеротрофної еукаріотичної клітини з ціанобактерією («синьо-зеленою водорістю») призвів до появи рослин, оскільки симбіотична ціанобактерія надалі трансформувалась у пластиду.
Синтетична гіпотеза ред.


Численні перевірки ендосимбіотичної гіпотези, виконані з широким використанням електронної мікроскопії, мікрохімічних та молекулярно-генетичних методів, з одного боку, підтвердили гіпотезу ендосимбіотичного походження мітохондрій та пластид, з іншого — автогенетичну гіпотезу походження ядра та більшості одномембранних клітинних органел[6]. Таким чином, на початку XXI ст. теорія походження еукаріотів сформувалась на основі синтезу обох гіпотез — як автогенетичної, так і ендосимбіотичної, і отримала назву синтетичної гіпотези походження еукаріотів.
Сьогодні вважають, що першим кроком на шляху виникнення еукаріотів було утворення у гіпотетичного прокаріотичного пращура численних внутрішніх інвагінацій плазмалеми, які, з одного боку, замкнули прокаріотичний нуклеоїд у двомембранну оболонку (тобто утворили морфологічно оформлене ядро), а з іншого — призвели до утворення ендоплазматичної сітки та похідного від неї комплексу Гольджі, а також травних вакуолей та їх похідних — лізосом.
На другому етапі, внаслідок горизонтального переносу гену, що кодує білок тубулін від спірохетоподібних бактерій, примітивні еукаріоти набули здатності до синтезу тубулінових мікротрубочок[3][7]. У результаті в еукаріотів з'явився цитоскелет, джгутики з базальним тілами, веретено поділу, мітоз. Базальні тіла джгутиків надалі у частини представників трансформувалися у клітинний центр, а порушення нормального мітозу (зокрема, скорочення інтерфази) призвели до виникнення мейозу і пов'язаного з ним статевого процесу. Група первинно безмітохондріальних еукаріотів сьогодні розглядається як перше, найпримітивніше царство еукаріотів — Hypochondria (Домітохондріальні, або Гіпохондріати)[8]. Всі гіпохондріати є одноклітинними гетеротрофами з тваринною стратегією живлення.
На третьому етапі еукаріотична клітина утворила симбіотичний комплекс із прокаріотичною клітиною, схожою із сучасними альфа-протеобактеріями. Ця прокаріотична клітина надалі трансформувалась у мітохондрію. За результатами як цитологічних, так і молекулярних досліджень, найдавнішими мітохондріальними еукаріотами вважаються ті, які мають мітохондрії з особливими дископодібними кристами. Група мітохондріальних еукаріотів з дископодібними кристами прийнята як таксон рангу царства — Discicristata (Дискокристати). В основі царства дискокристат перебувають первинно гетеротрофні організми. На четвертому етапі еукаріоти розділилися на дві великі групи. Одна з цих груп має мітохондрії з трубчастими кристами, і складає царство Tubulocristates (Тубулокристати, трубчастокристні), друга — мітохондрії з переважно пластинчастими кристами, і приймається як царство Platycrsitates (платикристати, платівчастокристні). Подібно до дискокристат, в основі філ тубуло- та платикристат знаходяться первинно гетеротрофні організми із тваринною стратегією живлення.
На п'ятому етапі в еукаріотичному світі з'явилися перші рослини. За даними молекулярної та клітинної біології, ця подія пов'язана з симбіозом гетеротрофного еукаріота-платикристати з фотоавтотрофним прокаріотом — ціанобактерією (синьо-зеленою водорістю). Як наслідок цього симбіозу утворилась пластида, оточена двома мембранами, яка отримала назву первинно симбіотичної пластиди. Подальша дивергенція організмів із первинно симбіотичними пластидами обумовила виникнення у межах філи платикристат групи фотоавтотрофних відділів, які склали підцарство Plantae — рослини. Дивергенція гетеротрофних платикристат за типами живлення зумовила виникнення з одного боку, відділів з осмотрофним живленням — підцарства Fungi (гриби), з іншого — таксонів з фаготрофним живленням, що склали підцарство Animalia — тварини.
Велика радіація тубулокристат відбувалась майже паралельно з платикристатами, проте була пов'язана не стільки з еволюцією за типами живлення, скільки з еволюцією клітинних покривів та джгутикового апарату. В основі тубулокристат залишились амебоїдні організми, які складають підцарство Ameboflagellates (амебо-флогеляти). Дві прогресивніші філи представляють, з одного боку, таксони зі специфічними, альвеольованими покривами — Alveolata (альвеоляти), та з іншого — зі специфічними субмікроскопічними тричленними.
Первинні та вторинні ендосимбіози ред.
Первинні ендосимбіози ред.

Ендосимбіози, при яких в еукаріотичну клітину-хазяїна включався прокаріотичний симбіонт, що надалі трансформувався в клітинну органелу, називають первинними. На основі молекулярно-таксономічних досліджень наприкінці ХХ ст. було встановлено, що у світі еукаріотів первинні симбіози відбулися лише двічі. При першому ендосимбіозі прокаріотична бактерія, схожа з альфа-протеобактеріями, вступила в симбіоз із фаготрофним джгутиконосцем з царства гіпохондріат і трансформувалась у мітохондрію з трьома типами крист — дископодібними, трубчастими та пластинчастими. Подальша морфологічна еволюція мітохондрії відбувалася поступово, майже паралельно з клітиною-господарем. Так, у тубулокристат дископодібні кристи були редуковані, а збереглися трубчасті та інколи разом з ними — пластинчасті кристи. У платикристат були втрачені як дископодібні, так і трубчасті кристи, а збереглися лише пластинчасті (за винятком відділу Haptophyta).
Інший первинний ендосимбіоз відбувся між прокаріотичним оксигенним фотоавтотрофом — ціанобактерією (синьо-зеленою водорістю) та гетеротрофним організмом із царства платикристат. Як наслідок, виникла перша еукаріотична клітина, здатна до оксигенного фотосинтезу — так звана «протоводорість», тобто утворились рослини. У «протоводорості» пластида була вкрита двома мембранами, з яких внутрішня являла собою мембрану прокаріотичного ендосимбіонту — ціанобактерії (синьо-зеленої водорості), а зовнішня була похідною від плазмалеми клітини-господаря, і ймовірно являла собою видозмінену мембрану травної вакуолі, в яку потрапив ендосимбіонт.
«Протоводорість» дала початок трьом паралельним гілкам рослин із первинно симбіотичними пластидами — глаукоцистофітовим водоростям (Glaucocystophyta), червоним водоростям (Rhodophyta) та зеленим водоростям (Chlorophyta). Усі ці три відділи зберігають пластиди, що оточені лише двомембранною оболонкою. Глаукоцистофітові водорості зберігають цікаву атавістичну ознаку — між зовнішньою та внутрішньою мембраною пластиди розташовується шар муреїну — речовини, характерної для клітинних оболонок більшості евбактерій, і, зокрема, синьо-зелених водоростей. Відділ глаукоцистофітових є сліпою гілкою еволюції рослин[джерело?].
Пластиди червоних водоростей — родопласти — також зберігають деякі яскраві ознаки спорідненості із «синьо-зеленими водоростями» (ціанобактеріями), зокрема, так звані фікобілінові пігменти. Пластиди зелених водоростей — хлоропласти — зберігають найменше фенотипічних рис схожості із прокаріотичним ендосимбіонтом. Від зелених водоростей беруть початок наземні рослини, причому всі вони зберігають первинно симбіотичні хлоропласти.
Вторинні ендосимбіози ред.
Червоні та зелені водорості неодноразово ставали ендосимбіонтами гетеротрофних еукаріотів із різних царств — дискокристат, тубулокристат та платикристат. Симбіози, що відбувалися за сценарієм «еукаріотичний хазяїн + еукаріотичний ендосимбіонт» називають вторинними. Внаслідок вторинних ендосимбіозів виникла велика група відділів водоростей із вторинно симбіотичними пластидами. Вторинно симбіотичні пластиди, на відміну від первинно симбіотичних, вкриті переважно чотирма мембранами, з яких внутрішня мембрана є залишком плазмалеми синьо-зеленої водорості (первинного ендосимбіонту), друга мембрана — видозміненою мембраною травної вакуолі господаря первинного ендосимбіонта (червоної або зеленої водорості), який є водночас вторинним ендосимбіонтом, третя мембрана — власне плазмалемою вторинного ендосимбіонта, четверта, зовнішня мембрана — плазмалемою господаря вторинного ендосимбіонта.
Цитоплазма, яка перебуває у проміжку між другою та третьою мембраною, так званий перипластидний простір, є редукованою цитоплазмою вторинного ендосимбіонта, і у деяких випадках зберігає 80S рибосоми та редуковану ендоплазматичну сітку. Зовнішні (третю та четверту) мембрани вторинно симбіотичних пластид називають хлоропластною ендоплазматичною сіткою.
Проте особливо цікавим є той факт, що у двох відділах водоростей із вторинно симбіотичними пластидами — хлорарахніофітових (Chlorarachniophyta) та криптофітових (Cryptophyta) — у перипластидному просторі зберігається навіть редуковане ядро вторинного ендосимбіонта — нуклеоморф. Нуклеоморф оточений двомембранною оболонкою і містить ДНК, яка (за наявними в нуклеоморфі генами) у хлорарахніофітових споріднена з ядерною ДНК зелених водоростей, а у криптофітових — із ядерною ДНК червоних водоростей.
У деяких водоростей, зокрема, в евгленових, вторинно симбіотичні пластиди вкриті не чотирма, а лише трьома мембранами. Вважається, що у цих водоростей одна з зовнішніх мембран пластиди була вторинно втрачена. Відомі випадки, коли пластиди (як первинно, так і вторинно симбіотичні) редукувалися, і тоді виникали вторинно гетеротрофні таксони. Зокрема, доведена вторинна редукція пластид у типу Apicomplexa, пращурами яких були фотоавтотрофні динофітові водорості (Dinophyta).
Є також дані, які дозволяють припустити, що оомікотові (Oomycota) утворились внаслідок вторинного переходу до гетеротрофного типу живлення від однієї з груп водоростей-тубулокристат. Крім того, вторинно гетеротрофні таксони низьких та середніх рангів (від видів до порядків і навіть класів) відомі майже в кожному відділі водоростей, а також серед вищих рослин[джерело?].
Поширення пластид через вторинні ендосимбіози обумовило виникнення значної кількості відділів водоростей. Наприклад, якщо водорості з первинно симбіотичними пластидами представлені лише трьома відділами (Glaucocystophyta, Rhodophyta, Chlorophyta), то водорості зі вторинно симбіотичними пластидами належать до 12 відділів. Серед них два відділи зберігають нуклеоморф (Chlorarachniophyta, Cryptophyta), 8 відділів мають чотиримембранні вторинно симбіотичні пластиди без нуклеоморфу, що походять від червоних водоростей (Raphidophyta, Chrysophyta, Eustigmatophyta, Xanthophyta, Phaeophyta, Bacillariophyta, Dictyochophyta, Haptophyta). Евгленофітові водорості мають тримембранні пластиди, пращурами яких є зелені водорості. У Dinophyta пластиди утворювались неодноразово, як внаслідок симбіозів із зеленими водоростями, так і навіть внаслідок третинних симбіозів із вторинно симбіотичними фотоавтотрофами.
Розмноження ред.
Поділ клітини відбувається безстатево мітозом, процес дозволяє кожному дочірньому ядру отримувати по одній копії кожної хромосоми. Цикл життя багатьох еукаріотів включає статеву репродукцію, змінюючись з гаплоїдної фази, де в кожній клітині наявна одна копія кожної хромосоми, на диплоїдну фазу, де в кожній клітині наявні по дві копії кожної хромосоми. Диплоїдна фаза являє собою злиття двох гаплоїдних гамет, що формують зиготу, котра в свою чергу може ділитись в мітозі або пройти хромосомне скорочення в мейозі. У цьому шаблоні допустимі значні варіації — у тварин відсутня багатоклітинна гаплоїдна фаза, проте кожне покоління рослин може проходити через гаплоїдну та диплоїдну багатоклітинну фази.
В еукаріотів відношення площі до об'єму менше, ніж у прокаріотів, тому метаболічний процес та процес утворення клітини триває довше.
Еволюція процесу статевої репродукції — це первісна та фундаментальна характеристика еукаріотичних організмів. Базуючись на філогентичному аналізі, молекулярні біологи Дакс та Роджер припустили[джерело?], що статеве розмноження, хоч і не було основним способом розмноження, все ж було притаманне спільному предку всіх еукаріотів. Основний комплект генів, котрі фігурують на стадії мейозу, наявний навіть у таких паразитичних видів як Trichomonas vaginalis та Giardia intestinalis, котрі донедавна вважали асексуальними. Оскільки ці два види є нащадками родів, що на ранній стадії відділились від головного еволюційного дерева еукаріотів, було припущено, що основні гени, які фігурують в фазі мейозу, а отже і статеве розмноження, були, швидше за все, притаманними спільному предку еукаріотів. Еукаріотичні види, що помилково вважалися безстатевими, наприклад, паразит з роду Leishmania, все ж мають статевий цикл. Також, певні чинники вказують на те, що амеба, котра вважалась безстатевим видом, з давніх часів розмножувалася статевим шляхом, та що в більшості сучасних безстатевих видів нещодавно та незалежно один від одного прокинувся сексуальний потяг[джерело?][сумнівно ].
Джерела ред.
- ↑ Словник української біологічної термінології / відпов. редактори: Д. М. Гродзинський, Л. О. Симоненко. — Київ : КММ, 2012. — 744 с. — ISBN 978-966-1673-12-9.
- ↑ Archibald, J.M., Cavalier-Smith, T., Maier, U. & Douglas, S. (2001). Molecular chaperones encoded by a reduced nucleus: the cryptomonad nucleomorph. J Mol Evol 52, 490501.
- ↑ а б в г Костіков І. Ю., Джаган В. В., Демченко Е. М., Бойко О. А., Бойко В. Р., Романенко П. О. Ботаніка. Водорості та гриби. — Київ, 2004.
- ↑ Thomas P. Curtis, William T. Sloan, and Jack W. Scannel (6). Estimating prokaryotic diversity and its limits. PNAS. 99 (16): 10494—10499. Архів оригіналу за 24 вересня 2015. Процитовано 19 серпня 2008.
- ↑ Retallack, G.J., Krull, E.S., Thackray, G.D., and Parkinson, D. (2013). Problematic urn-shaped fossils from a Paleoproterozoic (2.2 Ga) paleosol in South Africa. Precambrian Research. 235: 71—87. doi:10.1016/j.precamres.2013.05.015.
- ↑ Сергей Ястребов (22.11.2017). Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было. Новости науки на «Элементах». Архів оригіналу за 12 травня 2020. Процитовано 12 липня 2020.(рос.)
- ↑ P.J. Keeling, W.F. Doolittle Alpha-tubulin from early-diverging eukaryotic lineages and the evolution of the tubulin family. Molecular Biology and Evolution. 13: 1297—1305. doi:10.1093/oxfordjournals.molbev.a025576. PMID 8952074.
- ↑ Vossbrinck, C.R., J.V. Maddox, S. Friedman, B.A. Debrunner-Vossbrinck, and C.R. Woese (1987). Ribosomal RNA sequence suggests microsporidia are extremely ancient eukaryotes. Nature. 326: 411—414. doi:10.1038/326411a0. PMID 3550472..
Посилання ред.
- https://web.archive.org/web/20110825091526/http://shiva.msu.montana.edu/courses/mb437_537_2005_fall/docs/Cavalier-Smith.2002b.pdf
- https://web.archive.org/web/20081217144026/http://www.asm.org/ASM/files/ccLibraryFiles/Filename/000000003179/znw00707000335.pdf
- http://mbe.oxfordjournals.org/cgi/reprint/13/10/1297
- http://itol.embl.de/itol.cgi [Архівовано 8 липня 2007 у Wayback Machine.]
- http://www.ucmp.berkeley.edu/alllife/eukaryota.gif [Архівовано 13 липня 2007 у Wayback Machine.]
- ЕУКАРІОТИ [Архівовано 12 березня 2016 у Wayback Machine.] // Фармацевтична енциклопедія
{kind=link}
{kind=link}
| Це незавершена стаття з біології. Ви можете допомогти проєкту, виправивши або дописавши її. |